
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第49話
ホモ・サピエンスの進化
文と写真 長谷川政美
この連載もいよいよ終わりに近づいてきた。最後の3話でわれわれ自身の種であるヒトHomo sapiensの進化について触れておこう。◎チンパンジーからの分岐
1980年代までの動物園では、チンパンジーやゴリラの檻の前にはたいてい、「オランウータン科」と書かれた看板が立っていた。図49-1aのように、チンパンジー、ゴリラ、オランウータンは近い親戚と見なされていたのである。それに対して、現生霊長類のなかでヒトだけが唯一ヒト科に属すとされ、ヒトがオランウータン科と分かれて独自の進化の道を歩み始めたのは、3000万年以上前であったと考えられていた。
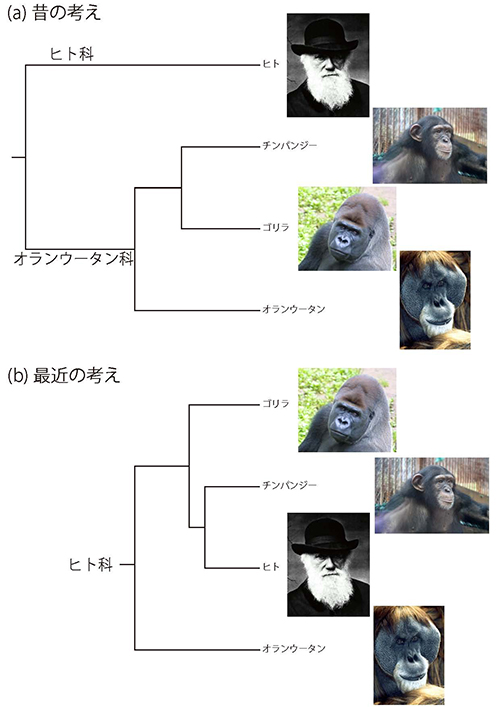
図49-1. ヒトに近い親戚に関する考え方の変遷。
この考えに対する挑戦は1962年頃から始まった。まだDNAの塩基配列を比較する現在の分子系統学が誕生する前のことであるが、その挑戦者は、アメリカのウェーン州立大学のモリス・グッドマン(図49-2)であった。

図49-2. モリス・グッドマンMorris Goodman(1925–2010)。1990年にニューヨークのコールド・スプリング・ハーバー研究所で開かれた進化に関するシンポジュウム会場にて。
彼の採った方法は、次のようなものであった。ヒトの血清中のアルブミンというたんぱく質をウサギに注射すると、ウサギの血清中に抗体ができる。この抗体を含む血清をヒトのアルブミンと混ぜると、強い抗原抗体反応が起って沈殿が生じる。次にこの血清をチンパンジーのアルブミンと混ぜると、ヒトとチンパンジーではアルブミンのアミノ酸配列が少し違うので反応は少し弱くなるが、やはり抗原抗体反応が起る。このような抗原抗体反応の強さを測ることによって、動物の種間でどのくらいアルブミンのアミノ酸配列が違っているかがわかるのである。こうして測られたアルブミンの違いを「免疫学的距離」という。グッドマンがヒトとチンパンジー、ゴリラ、オランウータンのあいだの免疫学的距離を測ったところ、思いがけないことが明らかになった。
チンパンジーやゴリラなどのアフリカ類人猿とアジアのオランウータンのあいだの免疫学的距離に比べて、ヒトとアフリカ類人猿の免疫学的距離は半分近くしかなかったのである。このことから、グッドマンは、チンパンジーやゴリラはオランウータンよりもヒトに近いと結論した。
その後、1967年にはアメリカ・カリフォルニア大学バークレー校のビンセント・サリッチとアラン・ウィルソン(図49-3)は、分子時計の考えを使ってこれをさらに推し進め、ヒトがチンパンジーやゴリラなどのアフリカ類人猿と分かれて独自の進化の道を歩み始めたのは、今からわずか400万年前であったと主張した。

図49-3. アラン・ウィルソンAllan Wilson(1934 –1991)。筆者が1987年にバークレーの彼の研究室を訪れたときに、彼に連れて行ってもらった中華レストランにて。アランは人類進化に関する分子進化学の先駆者であったとともに、絶滅した生物のDNAを解析する古代DNA研究の先駆者でもあった。しかし、白血病のために、この分野のその後の発展を見ることなく、1991年に50歳代の若さで亡くなった。彼の夢はスヴァンテ・ペーボに引き継がれ、ネアンデルタール人ゲノム研究として実を結ぶことになる。
当時の技術ではチンパンジー、ゴリラ、ヒト3者の関係は不明であったが、1990年代に入って豊富なDNA配列データが得られるようになると、図49-1bのように、ゴリラよりもチンパンジーがヒトに近いことが明らかになってきた。またサリッチとウィルソンは、ヒトとチンパンジーの分岐をおよそ400万年前としたが、現在では500~700万年前と考えられている。ただし、この分岐についてはあとでもう一度触れることになる。
◎ホモ・サピエンスの最後の共通祖先
筆者がバークレーにアラン・ウィルソン(図49-3)を最初に訪ねた1987年頃、彼の研究室はのちに「ミトコンドリアのイヴ仮説」と呼ばれるようになる現生人類の進化に関する重要な論文を発表していた。彼の研究室のレベッカ・キャンらが世界各地のヒト(現生人類)のミトコンドリアDNAを解析した結果、図49-4のような系統樹を得ていた。ただし、図の一番上にあるネアンデルタール人は彼らの最初のデータにはなく、その後10年経ってからスヴァンテ・ペーボらが付け加えたものであり、そのことはあとで触れるが、ペーボはキャンの研究がなされた頃はウィルソン研究室の一員であった。
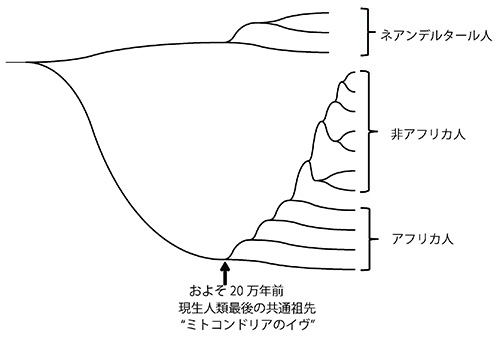
図49-4. 「ミトコンドリアのイヴ仮説」。
図49-4のネアンデルタール人を除いた部分の意味する重要なことは2点ある。まず、すべての現生人類のミトコンドリアの最後の共通祖先の生きていた年代がおよそ20万年前であること、次にそこから枝分かれするのは最初すべてアフリカ人の系統であり、非アフリカ人(ヨーロッパ人、アジア人、ニューギニア人、オーストラリア原住民、アメリカ先住民など)はもっとあとになってから枝分かれしていることである。
ミトコンドリアは組換えを起すことなく母系遺伝するので、祖先をたどっていくと当然一人の女性にたどり着くことになる。それが「ミトコンドリアのイヴ」であり、彼女は20万年前のアフリカで生きていて、現在76億を超える現生人類すべての人々にミトコンドリアを遺したのである。アフリカで進化した現生人類は、その後およそ10万年前にユーラシアに進出し、世界中に広がった。
◎ネアンデルタール人の発見
ダーウィンが『種の起源』を出版する3年前の1856年に、ドイツのネアンデル渓谷で古い人骨が発見された。これに関していろいろな議論があったが、現生人類とは遠い昔に分かれた系統だということで、ホモ・ネアンデルターレンシスHomo neanderthalensisという別種に分類されるようになった。
「旧人」と呼ばれるネアンデルタール人は、「新人」とも呼ばれる現生人類よりもがっしりした体つきで、脳も大きかった。彼らは、現生人類が10万年くらい前にユーラシアに進出した頃には、ヨーロッパや中東にいたが、その後およそ4万年前に絶滅した。彼らの絶滅の原因はよくわかっていない。
1997年に当時ドイツ・ミュンヘン大学にいたスヴァンテ・ペーボ(図49-5)らのグループは、1856年に発見されたネアンデルタール人の最初の標本からミトコンドリアDNAを解析することに成功した。

図49-5.スヴァンテ・ペーボSvante Pääbo(右)と筆者。1993年に彼が東京都港区にあった統計数理研究所の筆者の研究室を訪ねてきたときの写真。スヴァンテはその後、ネアンデルタール人のゲノム・プロジェクトを立ちあげて成功に導いた。
その後、ペーボは新しくライプツィヒに創設されたマックス・プランク進化人類学研究所に移り、ネアンデルタール人の古代DNA解析で画期的な成果を次々に発表していくことになるが、最初の数年間のターゲットはミトコンドリアDNAであった。その成果は、図49-4に示したようなものである。つまり、ミトコンドリアDNAで調べる限り、ネアンデルタール人は現生人類には何ら遺伝的な貢献をすることなく絶滅した種だということである。
以前は、人種の起源に関して、ヨーロッパ人はネアンデルタール人から進化し、東アジア人やオーストラリア原住民はホモ・エレクトスである北京原人やジャワ原人からそれぞれ進化したという考えがあったが、ペーボらの解析はこの考えを最終的に棄却したのである。
◎核ゲノム解析が明らかにしたこと
ミトコンドリアDNAのもつ情報は、遺伝情報のごく一部に過ぎない。2010年頃になると、ペーボのグループはネアンデルタール人の核ゲノムの解析に挑戦し、ついに成功する。その結果、ネアンデルタール人と現生人類の関係は、図49-4が示すような単純なものではないことが明らかになってきた。
図49-4からわかるように、ミトコンドリアDNAはアフリカ人のなかの多様性が非アフリカ人のなかの多様性を大きく上回る。ところが、カリフォルニア大学バークレー校のラスムス・ニールセンは、ヒトの核ゲノムのなかで非アフリカ人に見られる多様性がアフリカ人よりも大きい領域を12カ所見つけていた。ニールセンはペーボらのグループに加わり、この12カ所の領域をネアンデルタール人のものと比べてみたところ、そのうちの10カ所がネアンデルタール人と一致した。この領域では、図49-6が示すようなことが成り立っているのだ。ペーボとニールセンらは、このことはネアンデルタール人と非アフリカ人の祖先とのあいだで交雑が起り、ネアンデルタール人のゲノムの一部が、非アフリカ人の集団に持ち込まれたことを示していると考えた。
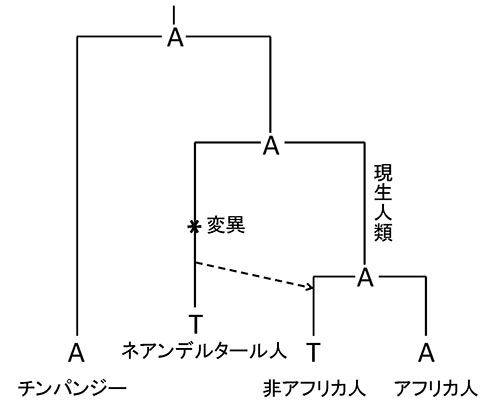
図49-6.ネアンデルタール人が非アフリカ人と交配したことを示す証拠。チンパンジーとアフリカ人の核ゲノムのある座位がA(アデニン)でネアンデルタール人と非アフリカ人でT(チミン)だとする。この場合、ヒトとチンパンジーの共通祖先でAだったものが、ネアンデルタール人の祖先の系統でTへの突然変異が起り、その変異が交雑を通じて点線矢印のように非アフリカ人に伝えられたと考えることができる。もちろん、交雑なしで非アフリカ人の祖先でもAからTへの変異が独立に起った可能性がある。しかし、このような座位が単独にあるのではなく、ゲノム中の特定の領域に集中して見られるならば、交雑が起ったと考えざるを得ないのである。デイヴィッド・ライク著『交雑する人類』(日向やよい訳、NHK出版)の図7を改変。
以上の解析で用いた現生人類のゲノムは現代人のものであるが、45,000年前のシベリアで生きていた現生人類のゲノムを調べてみると、ネアンデルタール人由来のDNAが現代の非アフリカ人のもつものよりも平均して7倍も長いことがわかった。つまり、この個体は、ネアンデルタール人との交雑が起って間もない頃のものであり、現代の非アフリカ人ではその後組換えが度重なったため、ネアンデルタール人由来のDNAは短くなっていったものと考えられるのだ。
このようなことから、図49-7が示すように、アフリカからユーラシアに進出した非アフリカ人の祖先が、ユーラシアに到達して間もなくネアンデルタール人と交雑して、ネアンデルタール人のゲノムの一部を取り込んだという考えが裏づけられた。
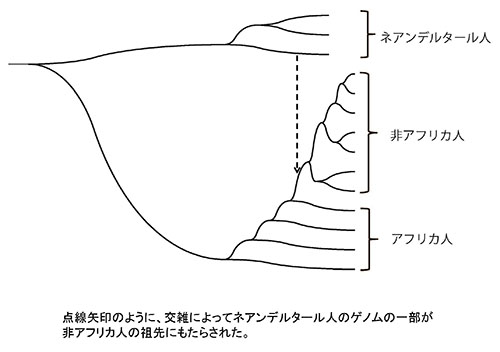
図49-7.ネアンデルタール人DNAの非アフリカ人への移入。
◎ネアンデルタール人由来のDNAを読み解く
非アフリカ人集団のなかのネアンデルタール人由来のDNAの割合は、平均2%だが、それはゲノム中に一様に分布しているわけではない。実はX染色体では少ないのだ。
図49-8は、AとBという2種が種分化するあたりの遺伝子の系図「遺伝子系統樹」を詳しく描いたものである。遺伝子の分岐は、必ず種分化の時点よりも古くなる。どれだけ古くなるかというと、平均して集団を構成する個体数に2を掛けた世代数だけ古くなるのだ。従って、2つの種が分化する少し前に、第3の種が分化していたとすると、遺伝子によってはこの3者のあいだの分岐の順番が図49-8の右下に示された種系統樹と食い違う関係を示すものが出てくる。
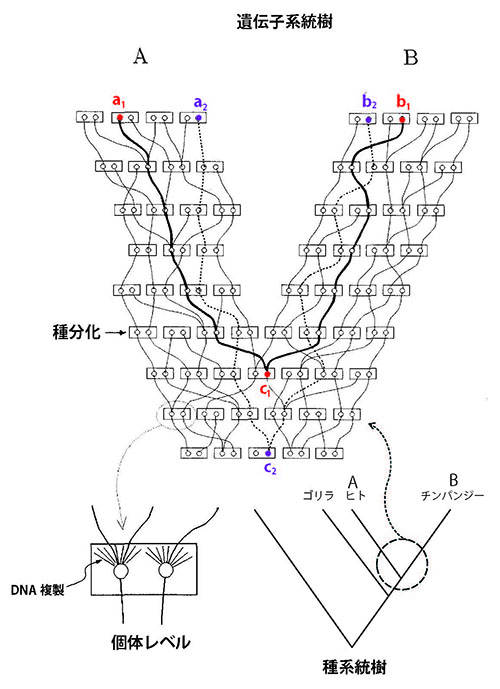
図49-8.AとBの2種が分岐するあたりを詳しく描いた「遺伝子系統樹」。長方形が個体を表わし、そのなかの2つの丸が父親と母親から受け継いだ遺伝子を表わす。種分化は、2つの集団のあいだの遺伝的な交流が途絶えた時点と考えておく。種Aから赤で示した遺伝子a1、種Bから遺伝子b1をサンプルしたとすると、これら2種の遺伝子の最後の共通祖先は種分化直前のc1になる。ところが、種Aから紫で示した遺伝子a2、種Bから遺伝子b2をサンプルしたとすると、これら2種の遺伝子の最後の共通祖先は、種分化よりも何世代か前のc2になる。このように2つの遺伝子の系統を遡っていってc1やc2のように合流することを「コアレッセンス」という。2種間の遺伝子のコアレッセンスは種分化の時点よりも必ず古くなるが、どのくらい古いかは集団の大きさに比例する。中立説が成り立つならば、集団の有効な大きさをNとすると、平均して2N世代だけ古くなることが知られている。ただしこれはあくまでも平均であり、ランダムに変動する。右下の種系統樹が示すように、Aがヒト、Bがチンパンジーだとすると、ヒトとチンパンジーが分かれるより前にゴリラが分かれているが、ゴリラの分岐はあまり古くないのでヒトとチンパンジーの遺伝子のコアレッセンスがゴリラの分岐以前まで遡る可能性がある。そのため、遺伝子系統樹は必ずしもヒト/チンパンジー近縁という種系統樹に一致するとは限らず、これとは違った関係を支持する遺伝子もある。Hasegawa, Kishino & Yano (1989)のFig.1を改変。
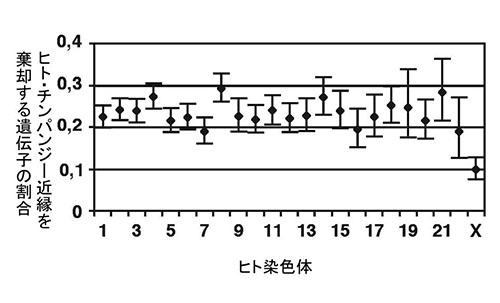
図49-9.ヒトとチンパンジーが近縁だという関係を棄却するような遺伝子系統樹の割合をその遺伝子が乗っている染色体ごとに表したもの。常染色体の遺伝子ではおよそ23%の遺伝子がヒト/チンパンジー近縁以外の関係を支持するが、X染色体ではそのような遺伝子はおよそ10%に過ぎない。性に関わらず2本ずつある常染色体に対して、X染色体はメスでは2本あるが、オスでは1本しかないため、X染色体の集団の大きさは、常染色体の3/4になる。そのために、図49-8のキャプションで説明したように種系統樹を棄却するような遺伝子系統樹が、X染色体では少なくなっていることが予想されるが、それだけではここで示されるほど少なくなることは説明できない。Ebersberger et al. (2007)のFig.3A。
X染色体の遺伝子系統樹がなぜこのように常染色体のものと違っているのだろうか。2つの集団が長く隔てられていると、次第に交雑で生まれる子供の生殖能力は低下する。このような生殖能力の低下は主にオスに起り、低下に関与する遺伝子はX染色体に集中している。ヒトとチンパンジーが500~700万年前に種分化した後しばらく経ってからこの2種のあいだで交雑が起り、そのとき生殖能力の低下をもたらす因子を排除する強力な自然選択が働いたと考えられる。X染色体にほかの種の遺伝子が残っていると生殖能力低下が起るために排除されたようなのである。
デイヴィッド・ライクによると、非アフリカ人集団のなかのネアンデルタール人由来のDNAの割合がX染色体では少ないということも、これと同じような機構で説明できるかもしれないという。
◎交雑を繰り返したヒト
核ゲノムの解析から、ヒトとチンパンジーや、現生人類とネアンデルタール人の関係は、かなり複雑だったことがわかってきた。いったん種分化したあとで、交雑によってゲノムが混ぜ合わされるということがしばしば起ったのである。さらに2018年の8月になって、ペーボらのグループは、ネアンデルタール人とそれとは別の旧人であり”第3の人類”とも呼ばれる「デニソワ人」とのあいだの雑種個体が5万年前のシベリア南部のアルタイ山脈で生きていた証拠を発表した。また、デニソワ人のゲノムの一部は、ニューギニア人やチベット人のゲノム中に見いだされている。チベット人の高地適応に、デニソワ人の遺伝子が寄与しているという研究もある。
系統樹が想定している単純な枝分かれは、大きな時間スケールで見た時の近似であり、枝分かれの近くを詳しく見ると、図49-10のようにいったん分かれて種分化が進んだあとで、枝が再び交わるような網目構造になっているのである。
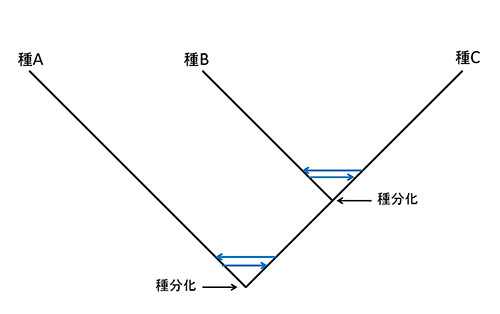
図49-10.交雑による網目状系統樹。青色の矢印は、種分化のあとの交雑による遺伝子移入を表わす。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


