
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第25話
エピジェネティックス
文と写真 長谷川政美
◎一卵性の双子の間の違い
一卵性の双子は全く同じゲノムDNAをもっているので、たいていは二卵性の双子よりもよく似ている。二卵性の双子のDNAは、普通の兄弟同士と同程度に違っているのだ。しかし、同じDNAをもっている一卵性の双子の間にも大きな違いが見られることがある。その原因の一つとして最近話題になっているのが「エピジェネティックス」である。
「エピジェネティックス」という言葉は、1942年にイギリスの生物学者コンラッド・ウォディントン(1905−1975)によってつくられたが、「後生説(エピジェネシス)」と「遺伝学(ジェネティックス)」を組み合わせたものである。
ウォディントンは、遺伝子と環境が相互作用することによって表現型が生み出されることを研究する新しい分野を、エピジェネティックスと呼んだのである。その当時、遺伝子の実体がDNAであることはまだ明らかになっていなかったが、現在ではエピジェネティックスとは、DNA塩基配列の変化によらずに受け継がれる遺伝子発現の変化(を研究する分野)とされている。
具体的には、DNAメチル化やヒストンの修飾などによって遺伝子発現が変化することを指すが、そのような変化が生殖細胞に及べば次世代に受け継がれる変異になり得る。このようなことが実際に起れば、進化的に大きな意味をもつことになる。次回紹介するが、確実に次世代に受け継がれるためには、受精したあとに起るエピジェネティックな変化の初期化を免れることも必要なのだが……。
2005年にスペイン国立がんセンターのマネル・エステラーらは、DNAメチル化とヒストンのアセチル化のレベルが一卵性の双子の間でどの程度違っているかを調べた。その結果、生まれたばかりの段階ではほとんど違いが見られないが、歳をとるにつれて違いが拡大することが分かった。特に、里子に出されたりなどして異なった環境で暮らした双子でその違いが顕著だった。
◎DNAメチル化による遺伝子発現の制御
DNAの塩基配列のなかで、シトシンCの次にグアニンGが現れるタイプの2連塩基をCpGという。2本鎖DNAで対になった両方のCpGのシトシンにメチル基が付加されることがDNAメチル化である(図25-1)。
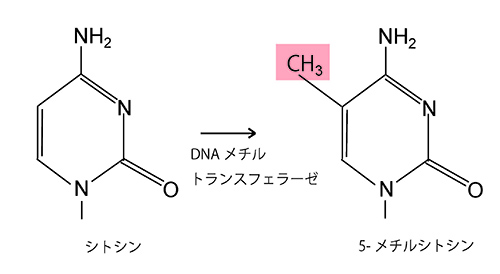
図25-1 DNAのメチル化。グアニンが続くシトシン塩基にメチル基(CH3)が付加されることによって起きるが、これには3種類のDNAメチルトランスフェラーゼという酵素が関与している。この図では簡単のために示されていないが、2つ以上の結合線が交差するところには炭素Cがある。
哺乳類では(たぶん脊椎動物一般で)、CpGという2連塩基は少ないが、その多くがたんぱく質をコードしている遺伝子の上流にある「プロモーター」と呼ばれる領域に集中している。プロモーターにRNAポリメラーゼが結合すると、たんぱく質をコードする遺伝情報のメッセンジャーRNA(正確にはその前駆体)への転写が始まる。ところがプロモーター中のCpGがメチル化されると、転写が抑制される。DNAメチル化により、遺伝子発現のスイッチがオフになるのだ。組織によって細胞内で発現する遺伝子は異なるが、DNAメチル化がこうした遺伝子発現を制御する仕組みの一つと考えられる。従って、分化した違う組織の細胞では遺伝子ごとのメチル化のパターンが異なっている。
細胞が分化するにつれてさまざまな遺伝子のプロモーターでメチル化が起きて、遺伝子発現のスイッチがオフになるが、細胞が分裂するときにそのメチル化パターンも複製されて受け継がれる。図25-2にその仕組みを示した。
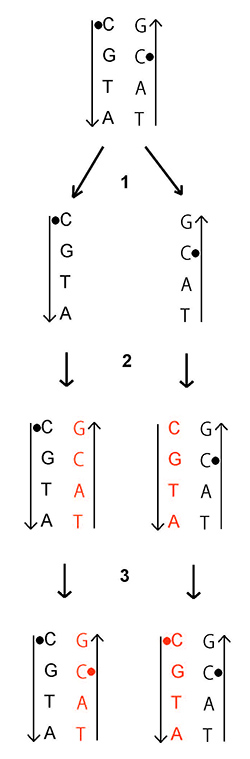
図25-2 DNAメチル化が複製される仕組み。DNAメチル化は、配列中で次にG(グアニン)が続くC(シトシン)に起る(図中●で示されている)。2本鎖DNAが複製される際は、まず1本鎖に分かれる(ステップ1).次にGに対してC、Cに対してG、A(アデニン)に対してT(チミン)、Tに対してAという規則で新しい鎖(オレンジ色)が合成される(ステップ2)。この段階では新しい鎖のCはまだメチル化されていない。次の段階で(ステップ3)、DNAメチルトランスフェラーゼ1という酵素が2本鎖の一方のCpGがメチル化されているのに、他方がメチル化されていないものを見つけ出し、そのようなCをメチル化する。このようにして、DNAメチル化も複製される。
哺乳類では、シトシンのメチル化が遺伝子発現の制御に深く関わっていることが明らかになっているが、DNAのメチル化はシトシンに限ったものではない。ショウジョウバエや線虫ではシトシンのメチル化はほとんど見られないが、最近になってこれらの動物でアデニンのメチル化が遺伝子発現の制御に関わっていることが示唆されている。
◎ヒストンアセチル化による遺伝子発現の制御
真核生物のDNAは、核のなかで折りたたまれたかたちで存在する。DNA(デオキシリボ核酸)は、五炭糖(デオキシリボース)とリン酸から成る2本の長い鎖がらせん状にのびた二重らせんである。この長い鎖をDNA主鎖という。2本の鎖は、DNA主鎖の糖に結びついている塩基対(アデニンとチミン、グアニンとシトシン)で結ばれている。主鎖のリン酸基は負電荷を帯びているので、そのままではたくさんの負電荷同士が反発しあうために、長いDNAをコンパクトに折りたたむことはできない。
なにしろヒトの1個の細胞に含まれるDNAの長さは2メートルにもなる。これが、直径が10マイクロメートル(0.01ミリメートル)ほどの核にしまい込まれているのだ。実際にはDNAは裸の状態にあるのではなく、正に帯電した塩基性たんぱく質であるヒストンに巻かれて存在している。リン酸のもつ負電荷を打ち消すように正電荷をもったヒストンがDNAを取り囲んでいるおかげで、DNAはコンパクトに折りたたまれるのだ。DNAとヒストンのこのような複合体をクロマチンという。
ただし、ヒストンがDNAにきつく結合したままでは、遺伝情報が発現するのに妨げになる。遺伝情報が発現するためには、RNAポリメラーゼがDNAに近づいてその情報をメッセンジャーRNAに転写することが必要なのに、ヒストンが邪魔になるのだ。
最近、生きた細胞内でクロマチンが揺らぐように動いている様子が観察されるようになった。つまり、ヒストンがいつもきつくDNAに結合しているわけではなく、揺らぐことによってRNAポリメラーゼのDNAへの接近を許しているのである。
このときに、ヒストンのアセチル化が重要な役割を果たしていることが分かってきた。ヒストンのなかのリシンというアミノ酸がアセチル化(図25-3)されることによって、その近くの遺伝子の転写が活性化されるのだ。
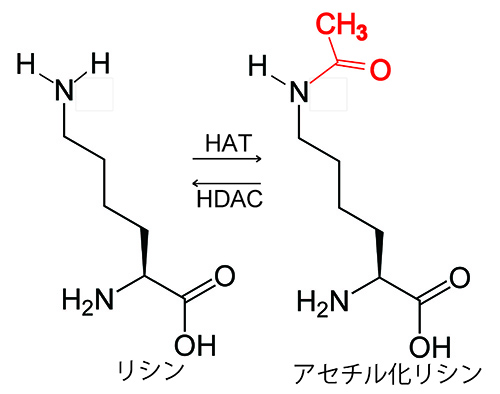
図25-3 ヒストンのアセチル化.ヒストンアセチルトランスフェラーゼ(HAT)がヒストンのリシン残基にアセチル基を付加することである.逆にヒストンデアセチラーゼ(HDAC)がアセチル化リシンからアセチル基を除去する反応がヒストンの脱アセチル化である。
図25-3では、リシンの側鎖は-NH2となっているが、通常は-NH3+の状態で存在する。つまり正の電荷を帯びているわけである。そのために、DNA主鎖のリン酸基の負電荷を打ち消すことで、ヒストンがDNAにきつく結合した状態が保たれる。ところがヒストンのアセチル化は、リシンの-NH3+の正電荷を相殺するために、ヒストンとDNAの結合が緩まり、遺伝子を転写するための因子がDNAにアクセスし易くなって転写が活性化されるものと考えられる。
ヒストンの修飾には、リシンのアセチル化以外にもさまざまなものがあり、遺伝子発現を活性化するものだけではなく、逆に抑制するものもある。
◎三毛猫の模様
哺乳類では、オスはX染色体とY染色体という2種類の性染色体をもっているのに対して、メスはX染色体だけを2本もっている。メスのもつ2本の染色体のうちの1本は不活性化して、そこにのっている遺伝子は発現しないようになっている。1本のX染色体で遺伝子発現が抑制されることをX染色体の不活性化という。1959年に大野乾(1928ー2000)は、哺乳類の雌の2つのX染色体のうちの1つは常染色体と同じように見えるが、もう1方は凝集している(バー小体という)ことを見つけた。この凝集した染色体が、不活性化したX染色体である。ヒストンの脱アセチル化やDNAのメチル化などによって、X染色体の不活性が起っているのだ。
メスのもつ2本のX染色体は、それぞれ父親由来と母親由来のものであるが、哺乳類のなかでカンガルーなどの有袋類では、父親由来のX染色体が選択的に不活性化される。ところが、ヒトやネコを含む真獣類では胚発生時にそれぞれの細胞で不活性化されるX染色体がランダムに決定され、その後細胞が分裂する際にもその不活性化状態が引き継がれる。
三毛猫の模様の形成には、このことが関係している(図25-4)。

図25-4 三毛猫。普通はメスネコで見られる。毛色を茶と黒にする遺伝子はX染色体上にあり、茶と黒が対立遺伝子である。父親由来と母親由来のX染色体上の遺伝子が、それぞれ茶と黒だと、三毛猫になる。メスネコの胚が100個ほどの細胞をもつ時期に、それぞれの細胞でどちらか一方のX染色体が不活性化され、分裂後もその状態が保持される。不活性化された遺伝子が茶であれば、その細胞は黒の色素を作り、逆ならば茶になる。そのために、このようなまだら模様が生まれる。
三毛猫は普通メスであり、白地に茶と黒がまだら模様になっている。白地の遺伝子は常染色体上にあるが、毛色を茶と黒にする遺伝子はX染色体上にあり、茶と黒が対立遺伝子の関係にある。これらの色素はメラニン細胞と呼ばれる細胞で発現する。メスネコの2本のX染色体上の遺伝子の一方が茶色、他方が黒色を指定していると、そのネコは三毛猫になる。
胚発生の過程で100個ほどの細胞をもつ時期に、メラニン細胞になるそれぞれの細胞でどちらか一方のX染色体がランダムに不活性化され、分裂後もその状態が保持されるのだ。そのためにまだら模様が生まれる。
将来メラニン細胞になるたくさんの細胞で、それぞれランダムに不活性化する染色体が選ばれるので、組み合わせの数は非常に多く、二匹の三毛猫の模様が同じくなることはない。一卵性双生児の三毛猫でも違う模様になる。また次節で触れる核移植によるクローン技術によって生まれた三毛猫の模様は、もとになったネコの模様とは全く違うものであった。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


