
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第22話
ジャンクDNA
文と写真 長谷川政美
◎ジャンクDNAはがらくたか?
21世紀に入り、多くの生物のゲノムの塩基配列が解読されるようになってきた。その結果分かったことは、ゲノムのなかでたんぱく質のアミノ酸配列をコードしているのはごくわずかの部分に過ぎないということであった。
ヒトのゲノムはおよそ30億塩基対から成るが、たんぱく質をコードする遺伝子の数は22,000個程度だと推定されている。1個の遺伝子のアミノ酸コード領域の長さが3,000塩基対(1,000アミノ酸)だとすると、ゲノム全体でアミノ酸コード領域が占める割合は、わずか2.2%に過ぎないのだ。残りの部分は何をしているのだろうか?
最初これらの大部分は「ジャンクDNA(がらくたDNA)」と呼ばれた。ところが、ヒトのゲノムを50から100塩基程度の短い断片ごとにマウスのゲノムと比較してみると、およそ5%の断片で配列が比較的よく保存されていることが分かった。つまりアミノ酸コード領域以外のジャンクDNAと呼ばれた部分でも、5%近くのDNA配列で負の選択圧が働いているようなのだ。
突然変異はゲノム全体でほぼ均等に起るので、配列が保存されているということは、それらのDNA配列にも機能的な制約があって、負の自然選択が働いていることを意味する。このことはこれらの配列が機能をもっていることを示唆する。第8話で述べたように、重要な遺伝子ほど突然変異全体のうちで中立的なものの割合が少なく、それだけ変わりにくいのだ。
たんぱく質をコードする遺伝子の数は、ヒトで22,000個程度と推定されていると述べたが、マウスもヒトと同じ程度の数の遺伝子をもっている。一方、ショウジョウバエでは14,000個、酵母で6,000個、大腸菌で4,000個とされている。ヒトのもっている遺伝子の数は、単細胞の大腸菌のせいぜい6倍程度でしかないのだ。さらに多細胞動物のなかでもわずか1,000個ほどの細胞しかもたないカエノラブディティス・エレガンス Caenorhabditis elegansという線虫は、ヒトとほとんど変わらない20,200個の遺伝子をもっている。
これらの生物の複雑性の違いを定量的に測ることは難しいが、ヒトでは予想されていたよりも少ない数の遺伝子で複雑な体制を作り上げているのだ。このことは、複雑な体制を作り上げるためには、新しい遺伝子を作るよりも、既存の遺伝子を使い方でさまざまに組み合わせる制御機構が重要であることを示唆する。
ジャンクDNAと呼ばれた配列にもかなりの部分に負の自然選択が働いていることから、それらの部分が遺伝子発現の制御に関わっているものと考えられる。
また、生物種間の違いは、遺伝子の違いによるよりも、同じような遺伝子が違ったパターンで使われることによることが多いと考えられる。わずか100個の遺伝子のスイッチのオン・オフでも、2100≒1.3 x 1030という膨大な数のパターンが可能なのである。
受精卵が発生の過程で多様な組織の細胞に分化するときだけでなく、種分化により多様な種が生み出される際にも、このような遺伝子発現スイッチのオン・オフのパターンの多様性が重要な役割を果たしているのではなかろうか。
◎非コードRNA
最近、これまでアミノ酸配列をコードしないジャンクDNAと呼ばれていたDNA配列のなかに機能的に重要な役割を果たしている配列の存在が浮かび上がってきた。
「非コードRNA」をコードするDNAである。「非コード」とはアミノ酸配列をコードしていないという意味である。もちろん、以前からリボソームRNAや転移RNAなどをコードするDNAや、転写開始領域(プロモーター)や調節領域(エンハンサー、転写を高める)などたんぱく質をコードする遺伝子の発現を調節するDNA領域は知られていた。しかし、新たに見つかった非コードRNAも遺伝子発現の制御に関与していることが分かってきたのである。ある種の非コードRNAは、ヒストンに修飾を付加することによって、遺伝子発現の制御に関与する。
先ほど機能的に重要な役割を果たしている遺伝子の塩基配列は、例えばマウスとヒトの間で保存されていると述べた。ところが、機能的に重要な役割を果たしていると考えられる非コードRNAのなかには、配列があまり保存的でないものも多い。このことから、これらの配列は重要でない、とは必ずしも言えない。むしろこれらがそれぞれの種で特異的な働きをしている可能性もあるのだ。
2005年に理化学研究所のグループが、マウスの転写産物について片っ端から配列決定を行なったところ、なんと23,000種類以上の非コードRNAが同定されたという。たんぱく質をコードする遺伝子の数と同程度なのだ。これらのRNAの大部分の機能はまだ不明であるが、遺伝子発現の制御に関与している多数の非コードRNAの存在が明らかになっている。機能が不明だったためにジャンクDNAと呼ばれた領域の多くで、何らかの機能が見いだされつつある。
さらに2012年には、ヒトゲノムのおよそ3/4の配列が、何らかの細胞で何らかの時期にRNAに転写されていることが明らかになった。ヒトの遺伝的病気の原因となるゲノム領域が特定された領域のおよそ90%は、いわゆるジャンクDNAのなかにあったという。
日進月歩のこの分野の現況を知りたいひとは、最近出版されたネッサー・キャリーの『ジャンクDNA』(中山潤一訳、丸善出版、2016年)や小林武彦さんの『DNAの98%は謎』(講談社ブルーバックス、2017年)を参照してほしい。
◎選択的スプライシング
話を続けよう。近年さらに、たんぱく質をコードする遺伝子が22,000個程度しかなくても、実質的にはもっと多くの遺伝子があるのと同じ働きを可能にする機構も見つかっている。選択的「スプライシング(Alternative splicing)」と呼ばれるものである。
真核生物の遺伝子の多くでは、DNAのレベルではアミノ酸をコードする領域が一つながりになっているわけではなく、途中で何カ所もアミノ酸をコードしない塩基配列が入り込んでいる。途中のアミノ酸をコードしない塩基配列をイントロン、アミノ酸をコードする塩基配列をエクソンという(図22-1)。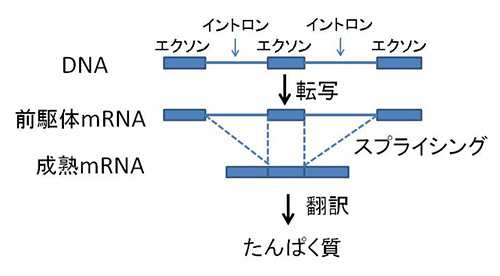
図22-1 イントロンとエクソン。
まずDNAからイントロンとエクソンを含む一つながりのメッセンジャーRNA(前駆体mRNA)が転写され、そのあとでエクソン部分だけがつなぎ合わされて(これをスプライシングという)成熟mRNAが出来上がる。
ヒトのダウン症にも関与しているといわれる「DSCAM(ダウン症候群細胞接着分子)遺伝子」と相同な遺伝子がショウジョウバエにもある。ショウジョウバエのこの遺伝子は、95個のエクソンから成るが、それらのエクソンは4つのタイプに分けられる。タイプ1、2、3、4にはそれぞれ12個、48個、33個、2個のエクソンが含まれる。前駆体mRNAがスプライシングされる際に、それぞれのタイプのエクソンから1個ずつが選ばれてつなぎ合わされるのだ。つまり一つのDSCAM遺伝子から、原理的には、12×48×33×2=38,016通りの違ったたんぱく質が作り出され得るのである(図22-2)。
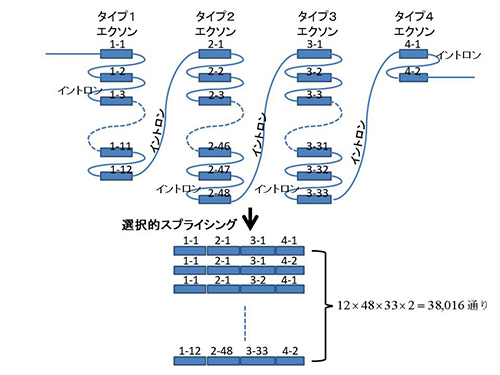
図22-2 DSCAM遺伝子における選択的スプライシング。
ヒトの遺伝子の少なくとも70%以上は、2種類以上のたんぱく質を作り出しているという。このように、ヒトのたんぱく質遺伝子の数が22,000個だとしても、実際に作り出されるたんぱく質の多様性はこれよりもはるかに高いのである。
◎たんぱく質の多機能性
複雑な生物のもつ遺伝子の数が予想よりも少ない理由として、1つのたんぱく質にいろいろな使われ方があるということも挙げられる。
脊椎動物の眼のレンズを作っているクリスタリンというたんぱく質には、いろいろな種類がある。そのなかで、ワニと多くの鳥類がもつε(イプシロン)-クリスタリンや鳥類と爬虫類だけで知られているδ(デルタ)-クリスタリンなど、特定の系統だけに見られるたんぱく質はすべて、もともとは代謝経路で機能する酵素であったという。眼が進化する過程で、透明な構造を作るのに都合の良い酵素を転用したものと思われるのである。この場合、酵素遺伝子の重複で酵素とは別に眼のクリスタリンが作られることもあるが、酵素としての機能をそのまま残したかたちで、レンズとして使われることもある。
多くの鳥がもつε-クリスタリンは、乳酸デヒドロゲナーゼという酵素そのものなのだ。また軟体動物の頭足類では、グルタチオン-S-トランスフェラーゼというまったく違った酵素をクリスタリンとして使うように進化した。
このように本来もっていた機能(本業)とは別の機能も併せて獲得した多機能のたんぱく質を「副業たんぱく質(moonlighting protein)」という。最近、たくさんの副業たんぱく質の存在が明らかになっている。進化とは、遠い将来のことも見越して綿密に計画された道に沿って進むのではなく、たまたま手に入る材料をやりくりして、その場をしのいでいくというかたちで進んできたのである。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


