
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第12話
ホッキョクグマの分布
文と写真 長谷川政美
◎なぜ白いのか
ホッキョクグマはその名前の通り、北極周辺の地域に分布する。彼らは厳しい寒さに適応したからだの構造をもっている。哺乳類の体毛はたいてい光を透過しないが、ホッキョクグマの体毛は光を透過し、内部が空洞の構造のために散乱される光によって白く見える。彼らの体毛は太陽光の通過を妨げないので皮膚にまで届き熱をもたらす。厚い脂肪層と内部が空洞になっている特殊な構造の体毛が、保温の役割を果たしている。図12-1の写真のように、彼らは泳ぎがうまく、氷の海を何時間も泳ぐことができるという。

◎ヒグマとの関係
進化的にはホッキョクグマはヒグマと非常に近縁である。ヒグマはユーラシアから北アメリカまで広く分布していて、形態的にも多様である。日本では北海道に生息し、エゾヒグマという亜種に分類される。
エゾヒグマは日本の陸上動物のなかでは最大であるが、ヒグマの中で最大のものは、アラスカ沿岸近くのコディアック島のコディアックヒグマで、雄は700kgを超えることもある。一方、チベットに生息するチベットヒグマ(ウマグマともいう)の体重は120kg程度しかない。このようにヒグマの大きさには地域ごとに大きな変異がある(図12-2)。
ホッキョクグマはそのなかで、特にアラスカ西海岸南端沖のABC諸島に生息するヒグマに近縁だという説がある。ヒグマの一地域集団からホッキョクグマが進化したというのだ。この問題はまだ決着がついていないが、少なくともヒグマの一地域集団とホッキョクグマの間で交雑を通じた遺伝的な交流は最近まで続いていたことは確からしい。

◎クマが南極に到達しなかった理由
いずれにしても、ホッキョクグマは北極圏周辺の地域でヒグマに似た祖先から寒冷地に適応した動物として進化した。いったん寒冷地適応した動物は、もはや熱帯の赤道を越えて南半球に進出することはできないので、南極に到達することは不可能であった。
およそ300万年前に、北アメリカ大陸は南アメリカ大陸と陸続きになり、メガネグマの祖先がクマとしては初めて南半球に進出した。6000万年前くらいまでは、南アメリカは南極と陸続きになっていたが、メガネグマの祖先が南アメリカに到達した頃には、南極大陸はすでに南アメリカからははるかに離れた孤立した大陸になっていて、メガネグマの祖先が到達することはなかった。
南極は孤立する以前は緑豊かな温暖な気候だったので、その頃までにヒグマのような動物が進出していれば、孤立して次第に寒冷化していく気候に適応して、ホッキョクグマのような動物が進化できた可能性はあったかもしれない。しかし、実際にはクマが進化したのはそれよりもずっとあとの時代だった。
南極ではホッキョクグマの代わりにヒョウアザラシが進化した。南氷洋にはウェッデルアザラシ、カニクイアザラシ、ロスアザラシ、ミナミゾウアザラシなどが魚、オキアミなどを食べるアザラシとして分布している。
南氷洋では、これらのアザラシやペンギンなどを捕食する動物としてヒョウアザラシが生態系の頂点にいる。北極海でアザラシを捕食しているホッキョクグマと同じ生態的役割を、南氷洋ではヒョウアザラシが果たしているのだ。
ペンギンがなぜ北極海にいないか、あるいはホッキョクグマがなぜ南極にいないか、といったことは、環境にあうように最適な種が創造されて配置されたのではなく、すべての種は種から生じたものであり、そこには移住が可能かどうかといった地理的な制約も重要であることを示している。そのような制約のもとでしか、進化は進行し得ないのだ。
◎アシカ科の起源
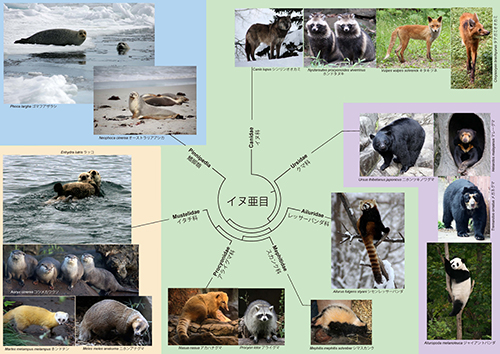
アシカやアザラシは鰭脚類と呼ばれ、食肉目のなかの1つのグループである。食肉目はイヌ亜目とネコ亜目に分けられる。図12-3にイヌ亜目の系統樹マンダラを示したが、鰭脚類はイタチ科、アライグマ科、スカンク科、レッサーパンダ科から成るイタチ上科と共通の祖先から進化した。
鰭脚類にはアザラシ科、セイウチ科、アシカ科が含まれる。このうちアシカ科は従来、アシカ亜科とオットセイ亜科に大きく分けられていた(図12-4)。

アシカ亜科には、北海道にも分布するトドや南アメリカのオタリアなどが含まれ、オットセイ亜科には北日本で見かけることがあるキタオットセイやニュージーランドオットセイなどが含まれる。
アシカ亜科では毛皮の下毛はまばらなのに対して、オットセイ亜科は密な下毛をもっている。しかし、私の研究室の大学院生だった米澤隆弘君がDNAの解析を行った結果、この分類は系統関係を反映したものではなく、南半球のいわゆる「オットセイ」は北半球のオットセイに近縁ではなく、それよりも同じ南半球に生息するアシカに近縁であることが明らかになった。鰭脚類全体の系統樹は図12-5のようになる。
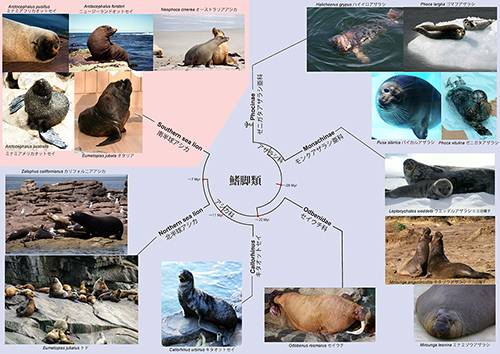
北半球のアシカ科には、キタオットセイ、トド、カリフォルニアアシカなどが含まれるが、絶滅したニホンアシカもこれに含まれる。一方、南半球には、オタリア、オーストラリアアシカ、ニュージーランドオットセイ、ミナミアメリカオットセイ、ミナミアフリカオットセイなど10種が分布している。現生種に関しては、南半球のほうが北半球よりも、種数と個体数のいずれに関してもはるかに多いのである。
ところが一方、化石記録に関しては、その大部分が北半球の北太平洋沿岸に偏っている。特にアシカ科の化石としては一番古い530~2300万年前の中新世のものは、北半球に限られ、南半球では180~530万年前の鮮新世になってようやく見つかっている状況である。つまり、現在のアシカ科の繁栄の中心は南半球だが、このグループの起源は北半球であったと考えられる。
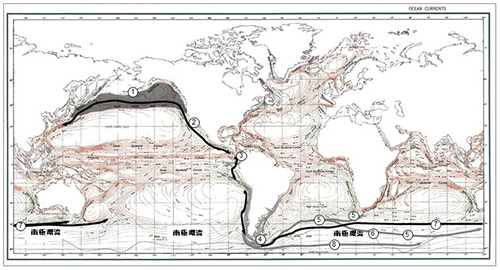
鰭脚類のなかでアシカ科に一番近いのはセイウチ科である。アシカ科がセイウチ科と分かれたのはおよそ2000万年前の中新世であったが、その後長い間、アシカ科の分布は北太平洋の図12-6の①で示された地域の沿岸に限られ、しかもほとんど多様化しなかった。
アシカ科が多様化しなかった原因は、同じ地域で多種多様なセイウチ科が繁栄していて、そのためにアシカ科の活動が制約されていたことにあったと考えられる。現在、図12-6の①で示された地域には、キタオットセイ、トド、カリフォルニアアシカが分布している。
現生のセイウチ科はセイウチ1種のみで、アシカ科とはあまり競合しない生活スタイルであるが、国立科学博物館の甲能直樹さんによると、その頃のセイウチ科の多くは現在のアシカ科と形態的によく似ており、食物や繁殖場所をめぐって激しい競争があったと思われるという。アシカ科は、中新世を通じてセイウチ科に圧迫されて、細々と生き続けていた。そのような状況だったとしたら、セイウチ科の分布しない南半球に移住すればよかったのにと考えられるが、それがそんなに簡単なことではなかったのだ。
◎移住の壁
そもそもアシカ科は寒冷な気候に適応に適応しているので、赤道を越えて南半球に移住することが非常に難しかったのである。ところが、中新世後期になってくると、気候が次第に寒冷化してくる。それにつれて、アシカ科も北アメリカ大陸の西海岸を伝って低緯度にも分布を広げるようになった(図12-6②)。
やがて670~580万年前になると、そのなかで赤道を越えて南半球に進出するものが現われたと考えられる(図12-6③)。その当時は、北アメリカ大陸と南アメリカ大陸はパナマ海峡で隔てられていたので、そこを通って大西洋に抜けるルートもあったが、そのルートは通らなかったようである。化石記録によると、当時のカリブ海沿岸にはセイウチ科がすでに進出していたからである。
南半球に進出したアシカ科は、南アメリカ沿岸に分布を広げた。図12-6④には現在オタリアとミナミアメリカオットセイが分布している。中新世後期の580万年前頃に、図12-6④あたりから、南極大陸の周りを西から東に向けて流れる南極環流に乗ってアフリカ大陸やオーストラリアへと分布を広げた。
その後も、何回か南極環流に乗った移住が繰り返された。また、南アメリカ大陸の西海岸を逆に北上し、ガラパゴスオットセイのように赤道近くのガラパゴスへ、さらにグアダルーペオットセイのように北緯29度に位置するメキシコ沖のグアダルーペ島にまで到達するものも現われた。
先にペンギンがガラパゴスよりも北に進出できなかったのは、冷たいペルー海流が西にそれるからだと述べたが、このことはそれと矛盾するようにも思われる。真相は分からないが、オットセイのほうがペンギンよりも暑さに強かったのか、あるいはペンギンもオットセイとともにグアダルーペまで到達したが、そこに定着できなかった事情があったのかもしれない。
いずれにしても、時間的なつながりと空間的なつながりを通じてはじめて進化の全貌が明らかになるのだ。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


