
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第46話
小さな生物が担う多様性
文と写真 長谷川政美
◎ニッチとフラクタル
マシュー・フィリップスとデビット・ペニーの新しい非鳥恐竜絶滅説は大変魅力的な仮説であるが、科学の世界では残念ながら魅力的な説が正しいとは限らない。まず問題になることは、非鳥恐竜の大きさの分布が、もともとほかの大部分の脊椎動物とずいぶん違っていることがある。
ネズミなどの齧歯目は哺乳類のなかでも小さなものが多いが、この仲間は哺乳類全体の種数のおよそ半数を占める。またスズメ目の鳥類も一般に小さいが、この仲間は鳥類全体の種数の60%を占める。このように小さな動物の種数が多い理由は、先に述べたように小さな動物ほど可能なニッチが多くなるからだと考えられる。
このことには、自然界がもっているフラクタル的な性質も関わっている。「フラクタル」とはアメリカの数学者ベノワ・マンデルブロが1960年代に発展させた数学の新しい概念である。図46-1は「コッホ曲線」と呼ばれるものであるが、図46-2にそれの描き方を示した。
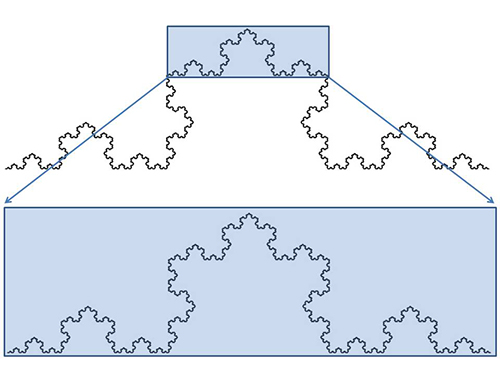
図46-1. コッホ曲線。1904年にスウェーデンの数学者ヘルジ・フォン・コッホが考案した曲線。部分が全体と相似的になっている。
。
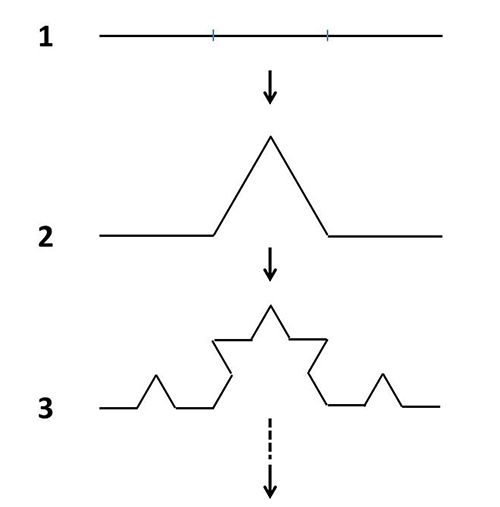
図46-2. コッホ曲線の描き方。①:まず線分を3等分する、②:続いて真ん中の線分を底辺とする正三角形を描き、底辺の線分を取り除く、③:②で残った4つの線分について、それぞれ1、2と同じ操作を繰り返す。④:③で残った16の線分について、同じ操作をする。このような操作を無限に繰り返して得られる図形が、図46-1のコッホ曲線である。線分の長さの合計は最終的に無限大になる。
コッホ曲線のような自己相似的な図形が「フラクタル図形」と呼ばれるものである。部分を拡大しても全体と同じようなかたちが見られるのだ。海岸線、川の分岐、血管の分岐、植物のかたちなど、自然界には近似的にフラクタルと見なすことのできるものが多い。
マンデルブロが1967年にサイエンス誌に発表した論文のタイトルは、「イギリスの海岸線の長さはどれだけか?」というものだった。海岸線の長さを実際に測ろうとすると奇妙なことが起る。普通の地図は海岸線のおおまかなかたちだけを描いていて、細かな凸凹は無視している。精度を上げて細かな凸凹も測るようにすると、大雑把な地図で測ったものよりも海岸線の長さは長くなる。精度をさらに上げていくと、海岸線の長さはどんどん長くなっていく。
海岸線がコッホ曲線だとすると、理想的なコッホ曲線は図46-2の操作を無限回繰り返して得られるものだから、線分の長さの合計は無限大になるのだ。これに対して、円周の長さを測る場合も精度を上げていくと次第に長くなるが、一定値(2πx円の半径)に収束する。
コッホ曲線などのフラクタル図形はこれとはまったく違う振る舞いをするのだ。マンデルブロのこの論文には「フラクタル」という言葉は使われていないが、8年後の1975年に彼がこの言葉を作り、その後、自然界のさまざまな現象がこの新しい言葉で記述されるようになった。
コッホ曲線は海岸線のように複雑ではあるが、完全に規則的なもので、個別の凸凹を特徴づけるものはない。一方、海岸線の凸凹は拡大しても似通って見えるが、完全な相似形ではなく、ランダム性ももち合わせている。個別の凸凹は、個別に特徴づけられた確率的なランダム性をもつのである。そのような違いがあるにもかかわらず、コッホ曲線は小さな動物ほど可能なニッチが多くなることを理解するには、よいモデルとなっている。
海岸線は拡大すると完全な自己相似的ではないが、いくら拡大しても似たような複雑な凸凹構造が残るということで、統計的に自己相似であると見なすことができる。このようなものを「統計的フラクタル」という。コッホ曲線を描く図46-2のやり方で、線分を3等分する際に乱数でばらつきを入れ、さらに真ん中の三角形を描く角度(60°、-60°など)にもばらつきを入れると、もっと現実の海岸線に近いものが得られる。マンデルブロ以前の古典的な数学の世界では、拡大していくと次第に直線に近づくが(これを微分可能性という)、現実の世界ではそうならないものが多いのである(とはいっても、微分学は自然科学にとって依然として重要なので、学生のあいだは学校ではきちんと勉強しよう)。
◎なぜ小さな生きものは種数が多いのか
3次元のコッホ曲線のようなものが動物の生息環境だとする。大きな動物にとっては、あまり変化に富んだ環境ではないが、小さな動物には入り込める隙間に富んだ環境である。凹んだ隙間と飛出た凸の部分では、全く異なる環境になる。理想的なフラクタル図形ならば、動物が小さくなればなるほど彼らにとっての環境の面積は際限なく広くなる。しかも、実際の生息環境では、例えばそれぞれの凸凹の太陽光に対する角度の違いなどで、さらに多様な環境が生まれる。
小さな動物は、大きな動物が認識できないような多様な環境の違いを感じ取りながら生きているのだ。従って、小さな動物ほど多くの種に分かれる傾向がある。しかも生息環境が広くなるので個体数も多くなる。小さな動物が感じている世界は、大きな動物が感じている世界よりも広いのである。これは単にからだの大きさに対して相対的に広いというのではなく、環境のもつフラクタル性から絶対的にも広くなっているのである。
このことは、100メートル四方、つまり1ヘクタールの樹木が数本生えた土地を考えてみると理解しやすいだろう。ルヤンゴサウルスのように全長38メートルもある巨大な恐竜にとっては、自分の餌となる植物が数本生えただけの文字通り1ヘクタールの狭い土地に過ぎない。ところが、体長10センチメートル程度のネズミにとっては、起伏に富んだ土地である。巨大な恐竜にとっては土地の凸凹は感じられなくても、ネズミにとってはその凸凹が隠れる場所を提供してくれたりする。さらにもっと小さな昆虫にとっては、樹の葉の裏表が異なる環境を提供するなど、小さな生物になるほど同じ1ヘクタールの狭い土地でも実際に利用できる彼らの生活の場が、面積で測っても広くなるのだ。
◎体重の分布
このようなわけで、哺乳類や鳥類の体重の種数分布は、図46-3の左に示したように、モード(最頻値)が平均値よりも左に偏った(右に裾をひいた)分布になる。横軸に体重の対数をとっても左に偏った分布になるのだ。
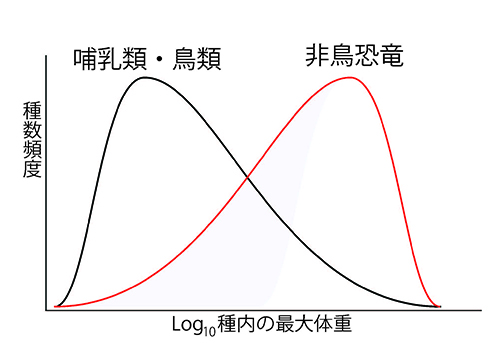
図46-3. 哺乳類・鳥類と非鳥恐竜の間の体重分布の比較。
このような分布が動物界全体で一般的であるが、そのことに環境のもつフラクタル性が関わっていると考えられる。フラクタル性だけからは、小さくなるほど動物の種数がどんどん増えていくことが予想されるが、実際には動物が生きていくための制約があって、そのようにはならない。偏りのない分布にくらべて左に偏った分布になる原因に、環境のもつフラクタル性が関わっているということである。
図46-4に、哺乳類のなかのネズミなどの齧歯目とコウモリなどの翼手目における実際の分布を示した。どちらにも左に偏った分布が見られる。ところが、非鳥恐竜の体重の種数分布は図46-3の右に示したように、脊椎動物に限らず動物一般のなかでは珍しく右に偏った分布なのである。
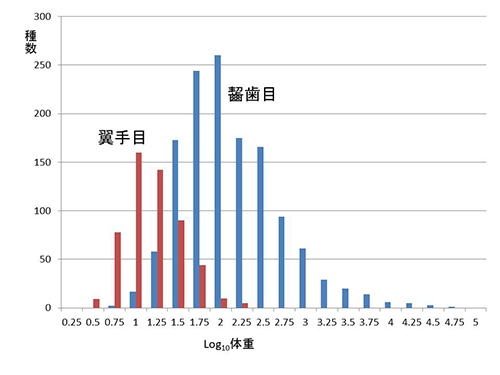
図46-4. 齧歯目と翼手目における体重の対数と種数頻度の関係。ただし、翼手目からはほかのグループと生態的に異なるオオコウモリは除いてある.データベースPanTHERIA http://esapubs.org/archive/ecol/E090/184/により作成。
非鳥恐竜の体重のこの奇妙な分布を生み出した原因は、グループ別に分布をみるともう少しはっきりしてくる。エオイン・オゴルマンとデビッド・ホーンは、非鳥恐竜を3大分類群、つまりトリケラトプスなどほとんどが植物食の鳥盤類、アパトサウルスやルヤンゴサウルスなど最大級の植物食恐竜を含む竜脚類、アロサウルスやなどほとんどが肉食恐竜の獣脚類に分けて、それぞれの体重の分布を調べた。その結果、植物食の鳥盤類や竜脚類では、体重分布が右に偏るのに対して、肉食の獣脚類ではあまり偏りのない正規分布に近い分布になることが分かった(図46-5)。
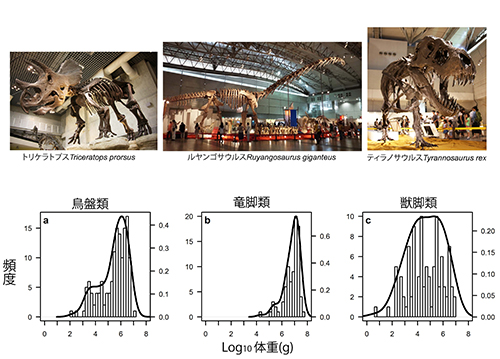
図46-5.非鳥恐竜の3大グループの間の体重分布の比較(O’Gorman and Hone, 2013)。トリケラトプスの写真は国立科学博物館、ルヤンゴサウルスとティラノサウルスの写真はギガ恐竜展(幕張メッセ, 2017)。
ところで従来は、竜脚類と獣脚類はあわせて竜盤類に分類されていたが、最近になって竜脚類は獣脚類よりも鳥盤類に近縁なのではないかという説が現れた。恐竜のこれら3大グループの間の系統関係には、まだ不明な点が残るが、いずれにしても、鳥盤類や竜脚類では肉食の獣脚類の捕食圧に対抗するためと、消化しにくい植物を効率よく消化するにもからだが大きくなるのが有利だということによって、体重分布が右に偏るようになったものと考えられる。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


