
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第38話
カンブリア爆発
文と写真 長谷川政美
◎ヒトにつながる動物の出現
カンブリア紀は5億4200万年前から始まる。当初この紀の始まりは、垂直に穴を掘った生痕化石が初めて現れる時期と定義された。そのような生痕化石は、運動性をもった動物が出現したということを意味する。しかし、前話で紹介したように最近ではそれよりも前のエディアカラ紀にすでに運動性をもった動物の痕跡が見いだされている。それでもわれわれに馴染みの動物が化石として現われるのはカンブリア紀になってからである。現存する30あまりの動物門にはっきりと該当する化石は、すべてカンブリア紀のあいだに現れた(図38-1)。






図38-1.およそ5億800万年前のカンブリア紀に現れたバージェス頁岩動物群(東京科学博物館2015年特別展「生命大躍進」ロイヤル・オンタリオ博物館所蔵)。(a)オパビニアOpabinia regalis(節足動物門)、(b)バージェソケタBurgessochaeta setigera(環形動物門)、(c)オットイアOttoia prolifica(鰓曳動物門)、(d)アイシェアイアAysheaia pedunculata(有爪動物門)、(e)ウィワクシアWiwaxia corrugata(軟体動物門)、(f)ピカイアPikaia gracilens(脊索動物門)。
これより遅れて現れた門はないと言われている。ただしなにごとにも例外があり、苔虫動物門はカンブリア紀の次のオルドビス紀(およそ4億5000万年前)に現れたという説があるが、カンブリア紀以降に新しい門がほとんど現れていないことは確かである。
この時期の化石として一番有名なのが、1909年にチャールズ・ウォルコットCharles Doolittle Walcott(1850-1927)がカナダ西部のバージェス頁岩のなかから発見した「バージェス動物群」であった。頁岩に閉じ込められた多種多様な動物のもので、節足動物、環形動物、軟体動物、脊索動物など明らかに現存する動物門に属するものが多く含まれていた。それらはおよそ5億500万年前のカンブリア紀中期の化石であった。それよりも古い化石のなかで現生の動物門と結びつけられるようなものは、ごくわずかしか発見されていなかったのだ。
その後、5億2500万年~5億500万年前の世界各地の地層からも同じような動物の化石が見つかった。なかでも中国雲南省の澄江(チェンジャン)で見つかるおよそ5億2500万年~5億2000万年前の「澄江動物群」が有名である。澄江動物群はバージェス動物群よりも古いが、よく似たものであるとともに、さらに広い内容を含んでいる。そのなかには、中国・西北大学の舒徳幹Shu Deganが2001年に新たな門として提唱した「古虫動物門ベチュリコラVetulicolia」の化石があるが(図38-2)、この門はその後絶滅した。
このように、およそ1500万年から2000万年という地質学的には短い期間に多様な動物門が化石として現われたということで「カンブリア爆発」と呼ばれている。

図38-2.ベチュリコラVetulicola cuneatus(古虫動物門、カンブリア紀前期、中国・雲南省・澄江Chengjiang、上海科学技術博物館所蔵)。この動物門はその後絶滅した。
創造論者にとっては、これは当然のことであった。カンブリア紀よりも古い時代の動物は知られていなかったので、カンブリア紀は創造主が生物を造られた時代だと見なされたのである。ダーウィンはそれに対して、将来カンブリア紀よりも古い時代の化石が見つかることに希望をつないだ。

図38-3.エオレドリキアEoredlichia intermedia(節足動物門・三葉虫類、カンブリア紀前期、中国・雲南省・澄江Chengjiang、上海科学技術博物館所蔵).三葉虫はこの後3億年にわたって続く古生代を生き延びた。
三葉虫はその後2億5000万年もの長い期間を生き抜くことになり、これまでに2万種近い三葉虫の化石が発見されているという。これは節足動物門に属するが、節足動物はバージェス頁岩動物群のなかで種数、化石数ともに一番多いグループである。現在でも節足動物は記載された種の数という点では断然トップの地位にあるが、カンブリア紀初期からその地位を保ち続けているのだ。
カンブリア紀の動物のなかで最もよく見られるのが三葉虫などの節足動物だが、そのほかのものもほとんどが背骨をもたない、いわゆる無脊椎動物である。それらのなかでピカイアという体長4センチほどの動物(図38-1f)は、脊索動物であり、このような動物からわれわれヒトを含む脊椎動物が進化したと考えられる。ピカイアは背骨の原型である脊索をもっていた。
◎動物界のなかの系統関係
アリストテレスの自然の階段では、動物は海綿動物、無血動物、有血動物の順に並べられていた(図1-1)。ラマルクは海綿動物と無血動物をあわせて無脊椎動物、有血動物を脊椎動物とし、動物界を2つのグループに分類した。ホヤやナメクジウオなども無脊椎動物とされた。「無脊椎動物」対「脊椎動物」という図式は最近まで踏襲された。
ところが、これが系統関係を反映していないことは、ダーウィンの時代から明らかであった。ホヤは英語名でascidianであるが、これはギリシャ語で「袋」を意味する。ホヤのからだを覆う被嚢が袋のようだからである。成体は海底の岩などに固着し、一見植物のようにも見える。からだの頂部に入水孔と出水孔という2つの管があり、一方から海水を体内に吸い込み、もう一方から排出する。この海水の流れによって、餌と酸素が取り込まれ、不要になったものが排泄される(図38-3のなかの右側にあるマボヤ参照)。成体はこのようなものであるが、ホヤの幼生は脊椎をもったオタマジャクシである。
ダーウィンは次のように述べている:
コワレフスキー氏から私が聞いたところでは、彼がその研究をもっと進めるならば、たいへん興味深い発見となるだろう。その発見というのは、ホヤの幼生はその発生の過程、神経系の相対的位置、そして脊椎動物の脊索に非常によく似た構造を備えているという点で、脊椎動物と近縁だというものである。……とほうもなく遠い昔、現在のホヤの幼生に多くの点で類似したグループの動物が存在し、それが二つの大きな枝に分岐し、その一つが発生の過程で後退して現在のホヤのなかまを生み出し、他方が脊椎動物を生み出すことによって、動物界の最高峰にまで登り詰めたと考えてよいだろう。
ダーウィン(1871)『人間の進化と性淘汰』
(長谷川眞理子訳、文一出版)
ダーウィンが正しく指摘しているように、脊椎動物はホヤの成体のような祖先から進化したのではなく、ホヤの幼生に似た祖先から進化したのである。ホヤも同じ祖先から進化し、成体が現在のように退化したかたちになった。オタマボヤというホヤは、その名前が示すようにおとなになってもオタマジャクシのままであり、これがホヤの祖先型と考えられる。現在では、ホヤはナメクジウオ、脊椎動物とともに、脊索動物門に分類される。
脊索動物門は棘皮動物門とともに「新口動物上門」に属する。「新口動物」とは、発生初期において原腸胚形成中に形成される陥入部の原口が口にならずに肛門となり、口は別に形成される動物のことで「後口動物」ともいう。新口動物の系統樹マンダラを図38-4に示した。
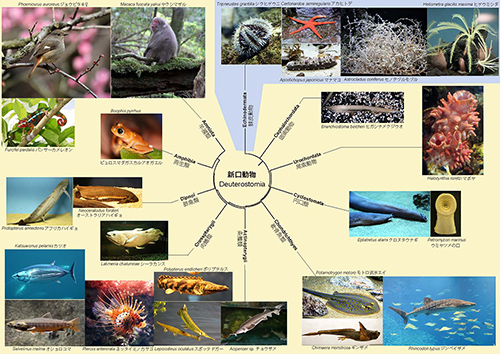
図38-4.新口動物の系統樹マンダラ。クリックすると大きく表示されます。
棘皮動物にはウミシダのように一見植物に見えるものもあり、ほかのものも左右相称動物とは言い難いように思われるが、棘皮動物の幼生は左右相称である。この図では、棘皮動物以外のものがすべて、脊索動物である。
脊索動物の共通祖先は、ナメクジウオやカンブリア紀のピカイア(図38-1f)に似たものだったと思われるが、そこからホヤと背骨をもった脊椎動物が進化した。最初の脊椎動物には顎がなく、図38-4の右に示されたウミヤツメの口のように円形の口をもっていたので、円口類と呼ばれる。そのなかから顎をもった魚類である軟骨魚類と硬骨魚類が進化した。
硬骨魚類のなかで現在普通に見られるのがすじ状の鰭をもった「条鰭類」であるが、その仲間から肉質の鰭をもったシーラカンスなどの「肉鰭類」や肺魚が進化した。肺魚との共通祖先から陸上脊椎動物(四足類)の「両生類」と「羊膜類」が進化した。
新口動物に対して、原口がそのまま口になるものが「旧口動物(前口動物)」である。前回、動物界全体の系統樹マンダラを図37-4で示したが、そのなかで「冠輪動物(外肛動物、扁形動物、腕足動物、軟体動物、環形動物)」と「脱皮動物(線形動物、有爪動物、節足動物)」が旧口動物である。以前は、ミミズなどの環形動物が同じように体節構造をもつ節足動物に近縁とされてきたが、分子系統学によって冠輪動物と脱皮動物という別の分類群に属することが明らかになった。
一方、線虫などの線形動物には体節がなく、節足動物との類縁関係はないと考えられていたが、同じ「脱皮動物」というグループに入ることがわかった。この名前が示すように、彼らは大きくなるときに表皮を脱ぎ捨てるのだ。脱皮動物という分類群の存在は、第32話でも登場したジム・レイク(図32-1)のグループによって1997年にはじめて指摘された。
新口動物と旧口動物をあわせたものが、三胚葉性の「左右相称動物」であり、これら活発に運動する動物は、大気中の酸素濃度がほぼ現在のレベルに達したエディアカラ紀以降にならないと出現できなかったものと思われる。
◎なぜ新しい動物門は生まれなかったのか
第20話で、モジュール要素の個性化によって動物の形態がさまざまに進化することが可能になり、そこでホメオティック遺伝子が重要な役割を果たしてきたことを紹介した。原生代の最後の頃までには、形態の進化をつかさどるそのような遺伝的な道具立ては揃っていたと思われる。
顕生代最初のカンブリア紀に入ると、カンブリア爆発と言われるように多様な動物門が化石として現れるようになる。ところが、それ以降、新たな動物門は現れないのである。それはなぜなのだろうか。
エディアカラ生物群は最後の全球凍結のあとで現れた。これには酸素濃度の上昇が重要な要因であったと思われるが、全球凍結によって多くの生物が絶滅し、競争相手のいない世界でエディアカラ生物群が進化したという側面もある。
カンブリア爆発でたくさんの動物門が一斉に出現したように見えるが、実際にはいつ動物門の分岐が起ったかについては、まだ不明な点が多い。しかし、地球の生態系がさまざまな役割を果たす動物で埋め尽くされる前に、分岐が起ったのは確かであろう。アンドルー・ノールは、「生物の多様化は、許容性の高い生態系が機能性の低い新種を生き残らせたときに開始する」と述べている。つまり、新しい動物門が出現するためには、最初は機能性の低いまったく新しいデザインを試してみる余地が必要である。資源をめぐる競争が過酷な状況では、そのような余地はあり得ないのだ。
カンブリア爆発以降は、資源をめぐる競争は厳しさを増し、新しい動物門が生まれる状況が訪れることはついになかった。ときには大量絶滅は起こり、そのつど生き残った生物からの多様化は起ったが、動物の基本的な体制が変わることはなかった。
現存の動物門とはっきり対応する化石が見つかるのはカンブリア紀に入ってからであるが、動物門のあいだの分岐の大部分はエディアカラ紀のあいだに起ったものと思われる。
およそ6億年の動物進化の歴史を通じて、基本的な体制である門のレベルの多様化が徐々に進んできたのではなく、エディアカラ紀からカンブリア紀にかけての、動物進化の歴史全体から見ると短い時間のあいだにすべての門が出揃ったのである。そのあいだに出揃った動物門のなかには古虫動物門のようにその後絶滅したものもあるが、そのほとんどは現在まで存続している。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


