
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第24話
ヘモグロビンにおける調節
文と写真 長谷川政美
ヘモグロビンというたんぱく質は、バクテリアからヒトに至るまでさまざまな生物で重要な働きを果たしている。バクテリアで見られるヘモグロビンには、細胞内の酸素など有害な分子を吸着して細胞外に排出する役割を負ったものもある。ヒトのヘモグロビンは、肺で取り込んだ酸素をからだの隅々まで運んで、活発なエネルギー代謝を支えている。
脊椎動物の系統においては、ヤツメウナギなど無顎類では、一本のポリペプチド鎖がヘモグロビンの役割を果たしているが、顎をもった有顎類(軟骨魚類、硬骨魚類、四足動物など)では2本のヘモグロビンα(アルファ)鎖と2本のヘモグロビンβ(ベータ)鎖の合計4本のポリペプチドが組み合わさって1つのヘモグロビン分子が出来上がっている。α鎖とβ鎖の遺伝子は、無顎類との共通祖先では1つであったものが有顎類の系統で重複して分化したものである。
4つのポリペプチド鎖には、それぞれ鉄原子1個を含むヘムがつながっていて、各鉄原子に酸素1分子が結合できる。血液中の赤血球の中にある4本鎖のヘモグロビンは、からだの隅々にまで酸素を運び、高度の運動性をもつ脊椎動物のエネルギー代謝を支えている。
四足動物でのヘモグロビンの役割は、肺で酸素を吸着し、それを組織に運んで、そこで放出することである。
放出した酸素と交換に、組織で二酸化炭素を吸着し、それを肺まで運んで放出する。肺には酸素が多く、組織には二酸化炭素が多いのだ。この機能を効率よく果たすために、ヘモグロビンは特別な性質を進化させた。図24-1に酸素濃度に対してヘモグロビンがどのくらいの割合で酸素を吸着するかを示した。
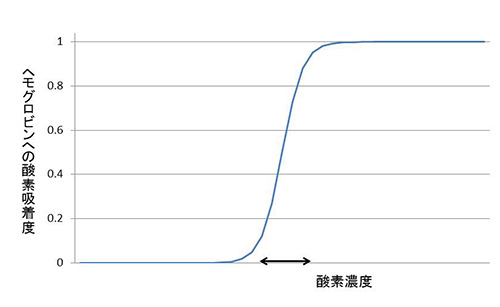
図24-1 ヘモグロビンの酸素吸着曲線。
ヘモグロビンは、2つの互いに違った立体構造をとることができる。一方の構造は酸素結合型、もう一方は酸素放出型である(図24-2)。
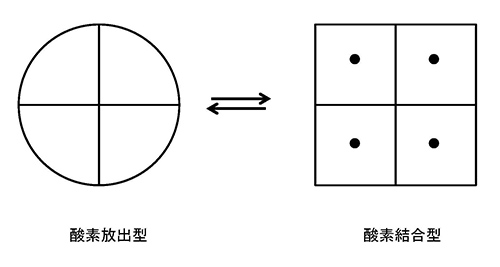
図24-2 ヘモグロビンの2つの状態を模式的に示した.黒い丸は酸素が吸着していることを示す。
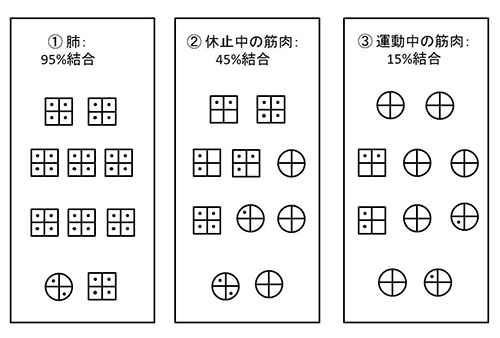
図24-3 四足動物におけるヘモグロビンの応答。①血液が肺を通過すると、ほとんどのヘモグロビン分子は酸素結合型になる。②血液が休止中の筋肉を通過すると、およそ半分のヘモグロビン分子は、酸素放出型になる。③血液が激しい運動中の筋肉を通過すると、大部分のヘモグロビン分子は酸素放出型になる。マーク・カーシュナー、ジョン・ゲルハルト著「ダーウィンのジレンマを解く:新規性の進化発生理論」(滋賀陽子訳、赤坂甲治監訳、みすず書房、2008年)の図を改変。
このようなことは、生命活動のまったく別の場面である遺伝子発現の制御に関わるレプレッサーなどのたんぱく質でもよく見られる。例えばある低分子がレプレッサーに結合すると、レプレッサーの離れた部位のDNAへの結合をゆるめて、遺伝子が発現しやすくなる。
ヘモグロビン分子に平均して2個の酸素分子が結合すると、酸素放出型と酸素結合型のヘモグロビンが半々ずつになる。逆に、4つの酸素を結合して酸素結合型にあったヘモグロビンのサブユニットの1つが酸素を放出すると、全体が酸素放出型に変わりやすくなる。
ヘモグロビンのもつこのような性質のため、図24-1の矢印の範囲で、少し酸素濃度が上がっただけで、雪崩打つように酸素がヘムに結合する。このような性質がなければ、肺から組織をめぐって肺に戻るまでの間で、結合している酸素のかなりの割合を放出することはできない。以上のような巧妙な仕組みは、図24-2で示したように、ヘモグロビンが2つの状態の間の平衡にあることからきている。このような平衡は、状況に応じて平衡をどちらかの方向に偏らせるように調節することも可能にする。
◎ヘモグロビンの調節
われわれが標高の高い高地に行くと、酸素濃度が低いので酸素を十分に取り込むことができなくなる。そのような状況では、血液中の2,3-ジホスホグリセリン酸という分子の濃度が上昇し、これがヘモグロビンの酸素結合の阻害剤として働いて、酸素親和性が低下する。酸素親和性の低下は肺でよりも組織内での効果が大きく、そのため組織内での酸素放出量を増やすことができるので、われわれはある程度までの低酸素濃度に耐えることができる。
図24-4にそのことを説明した。
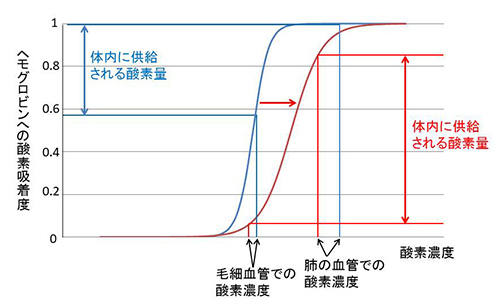
図24-4 ヘモグロビンの酸素吸着曲線。シグモイド曲線が右側に偏ると、体内に供給される酸素量が増える。
このようになると、低酸素状態になった肺で吸着する酸素量がさらに減るので一見不利に見える。ところが、ヘモグロビンが酸素放出型に偏っているために、毛細血管で放出しやすくなっているので、図の右側で示したように体内に供給される酸素量は増えるのである。
哺乳類では低酸素条件で取り込む酸素量を増やすために2,3-ジホスホグリセリン酸を使うが、鳥類ではイノシトール5リン酸を使うなどそれぞれの分類群でさまざまな分子を使うように進化した。
胎盤で育つ真獣類の胎児は、母親の肺から遠い場所にいるので、十分な酸素を得るために母親の組織と競合する。そのため、胎児の赤血球では母親の赤血球よりもヘモグロビンの酸素結合の阻害剤である2,3-ジホスホグリセリン酸の生産が少なくなっている。こうして、胎児のヘモグロビンは酸素結合型の状態に留まって、周辺の母親の組織を多少犠牲にして酸素を結合させることができる。
さらに真獣類のなかでも霊長類では、ヘモグロビンの遺伝子重複が起って、胎児用のヘモグロビンが進化した。ヒトの胎児ヘモグロビンは、成体のヘモグロビンとほぼ同じ酸素親和性をもつが、アミノ酸配列が数カ所違うため、2,3-ジホスホグリセリン酸と結合しにくく、酸素結合型に留まる傾向がある。
こうして母体から十分な酸素を得ることが出来るようになっている。このような胎児用のヘモグロビンは真獣類のいろいろな系統で独立に進化した。
このようにヘモグロビンの平衡状態を調節し、どちらかの方向に偏らせることは、ほかにもいろいろな方法で実現することができる。体温が上昇することもシグモイド曲線を右側に偏らせる。激しく運動をすると体内ではたくさん酸素が消費されるので、酸素が解離しやすい状態にあることが必要になるのだ。
先に標高が高く、酸素濃度の低い場所に行くと、血液中の2,3-ジホスホグリセリン酸という分子の濃度が上昇してシグモイド曲線が右側に偏るという話をした。実際には、低酸素状態に対応する仕組みはたくさん用意されており、さまざまな対応の仕方が可能である。
昔から高地に適応してきた人たちとしては、ヒマラヤ山脈の北側の高地に住むチベット人、南アメリカのアンデスに住むアンデス高地民、さらにアフリカのエチオピア高地民などがいるが、彼らの低酸素に対する適応の仕方は、それぞれに異なっている。
これらの人々は、低地にくらべて酸素濃度が2/3程度しかないような環境で暮らしてきたが、いずれもヘモグロビン分子そのものが変化しているわけではなく、何らかの調節機構の変更で対処していたのである。
しかしながら、鳥類の場合には、高地の低酸素濃度の環境への適応が、ヘモグロビン分子のアミノ酸配列を直接変えることによっても実現している例も知られている。インドガンAnser indicusは標高3000メールを越えるチベット高原で繁殖する(図24-5)。

図24-5 繁殖のためチベット高原にやってきたインドガンAnser indicus(中国青海省・青海湖にて、海抜3200メートル)。
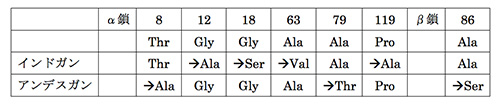
実は同じガンの仲間でやはり酸素濃度が低いアンデスの高地に住むアンデスガンChloephaga melanopteraがいる。この2種はガンの仲間のなかで特に近縁というわけではなく、それぞれ独自に低酸素の環境に適応したのである。表24-1に示したように、インドガンとアンデスガンのヘモグロビンのアミノ酸配列は、低地に住むほかのガンにくらべて変化しているが、それぞれに違ったアミノ酸置換で適応している。
表24-1 インドガンとアンデスガンにおけるヘモグロビンのアミノ酸置換.一番上の欄の数字はアミノ酸座位の番号、次の欄はインドガンとアンデスガン以外のガン亜科(ガン+ハクチョウ)で保存されているアミノ酸を示す。α鎖の8番目の座位でアンデスガンが “→Ala”となっているのは、アンデスガン以外のガン亜科はすべてThr(トレオニン)なので、共通祖先もThrであったものがアンデスガンの系統でAla(アラニン)に置換したと考えられることを示す(McCrackenら,2010より)。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


