
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第32話
真核生物の起源についての
「水素仮説」
文と写真 長谷川政美
◎古細菌と真正細菌の融合
2004年にアメリカのカリフォルニア大学ロスアンゼルス校のマリア・リヴェラとジム・レイク(図32-1)がゲノムの比較から面白いことを見つけた。
3大分類群のそれぞれからいくつかの代表を選んで全ゲノムを比較した結果、真核生物のゲノムが古細菌由来の遺伝子と真正細菌由来の遺伝子から構成されていることを明らかにしたのだ。
ここまでは、これまで述べてきたように、古細菌に近縁な真核生物の祖先に真正細菌のαプロテオバクテリアが共生し、共生体のゲノムが宿主の核ゲノムに転移したという話と同じである。

図32-1 ジム・レイク Jim Lake。1994年に著者がアメリカ・ロスアンゼルスの彼の自宅を訪ねたときの写真。
彼らは、真核生物の核の遺伝子を「情報系遺伝子」と「操作系遺伝子」に分類して調べたのだ。彼らのいう情報系遺伝子とは、DNAの複製や転写、たんぱく質合成などに関与する遺伝子であり、操作系遺伝子とは細胞の代謝、つまりエネルギー生成、脂質やアミノ酸の合成などに関わる遺伝子である。
彼らは、真核生物のほとんどの操作系遺伝子はαプロテオバクテリアに近縁な真正細菌由来であり、情報系遺伝子は古細菌由来であることを示した。
リヴェラとレイクは、真核生物が古細菌と真正細菌の融合によって生まれたのだと主張したのだ。確かに真核生物の遺伝子の由来をたどると、彼らが主張する融合説も有力な仮説の一つと考えられるが、まだ多くの疑問が残る。
最大の疑問は、真核生物の特徴である細胞核がどうして生まれたかという点である。カヴァリエ・スミスのアーケゾア仮説でも、まずアーケゾアという細胞核をもった祖先型真核生物が生まれ、プロテオバクテリアがそれに共生してミトコンドリアになったというシナリオであったが、細胞核の起源に関しては説得力のある説明はなかった。
◎メタン生成菌
近年、地球上の最初の生命は中央海嶺の熱水噴出孔の近くで生じたという説が有力である。西オーストラリア北部のピルバラで、35億年前の中央海嶺熱水系微生物化石の炭素同位体を解析した結果、古細菌のメタン生成菌と同じような炭素固定反応を行なっていた生物がいた証拠が見つかった。
第29話でも似たような方法を紹介したが、代謝の異なる生物では利用している炭素同位体の組成が違っている。微生物化石の炭素同位体組成を調べたところ、35億年前に現在のメタン生成菌と同じような代謝系をもった微生物がいたことが確認されたのだ。
これが現在のメタン生成菌と同じ起源をもつものか、独立に生じた系統かは不明であるが、このようなものが非常に早い時期に出現していたことは、次に紹介するミトコンドリアの起源に関する「水素仮説」にとって重要である。
◎ミトコンドリアの起源に関する「水素仮説」
先にトリコモナスというミトコンドリアをもたない真核生物がアーケゾアの候補として出てきたが、この生物はミトコンドリアをもたないが、その代わりに「ヒドロゲノソーム」という別の細胞内小器官をもっている。
ヒドロゲノソームは、ミトコンドリアと同じようにエネルギー生成に関わっているが、水素分子を老廃物として放出する。ヒドロゲノソームHydrogenosomeという名前は、水素Hydrogenを産生することからつけられたものである。ヒドロゲノソームはピルビン酸やリンゴ酸などを分解してATPを合成するが、そのときに水素と二酸化炭素を放出する。
前回登場したロックフェラー大学のミクロス・ミュラー(図31-4)は、長年このヒドロゲノソームを研究していた。ほとんどのヒドロゲノソームは自分自身のゲノムをもたないので、この細胞内小器官の起源は謎に包まれていた。しかし、ミュラーはこれがミトコンドリアと共通の祖先に由来するものではないか、と考えていたのだ。
1998年にドイツのビル・マーチン(図32-2)は、ミュラーと一緒に、「真核生物の起源に関する水素仮説」(以下では「水素仮説」と呼ぶ)というまったく新しい考えを発表した。図32-2の写真は、その2年後に彼らの「水素仮説」について講演してもらうために、著者が彼を日本進化学会に招待した際のものである。

図32-2 ビル・マーチンBill Martin(左)と著者。2000年に東京で開催された日本進化学会に彼を招待講演者として招いたときの写真。
マーチンとミュラーが注目したのは、「メタン生成菌」という嫌気的(酸素のない)な環境でメタンを作り出す古細菌であった。汚泥で泡がブクブク出ているのはこのメタン生成菌のせいであり、ウシの腸内にも棲んでメタンガスを発生させる。また天然ガスもほとんどがメタン生成菌によって作られている。
メタン生成菌は水素分子を二酸化炭素と反応させて、ブドウ糖(C6H12O6)などの生きていくために必要な有機化合物やエネルギーを作り出すことができるが、このときにメタンを放出する。酸素分子のある環境では、水素はすぐに酸素と反応して水になってしまうので(第29話:図29-5b)、メタン生成菌はそのような環境ではエネルギーを作り出せない。
一方、ヒドロゲノソームは、二酸化炭素と水素を放出するので、これがそばにいればメタン生成菌が生きていく上で必要な水素がそこから得られることになる。
マーチンとミュラーは、図32-3aが示すように、ブドウ糖などの有機物を分解してATPを合成し、そのときに二酸化炭素と水素を放出する、現在のヒドロゲノソームのような代謝を行う真正細菌がメタン生成菌の近くにいたと考えたのだ。
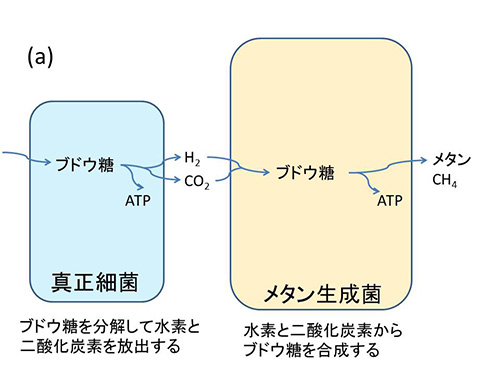
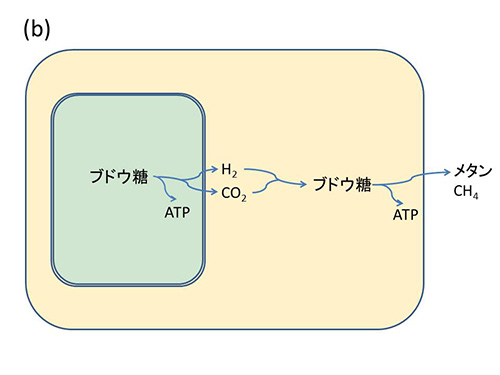
図32-3 ミトコンドリアの起源に関する水素仮説。
プロテオバクテリアは非常に多様な真正細菌の門であり、リケッチア、大腸菌、サルモネラ、ビブリオなど病原菌も含まれるが、窒素固定に関わる細菌など、自由生活性のものも多く含まれている。プロテオバクテリアという名前は、その多様性から、ギリシャ神話で姿を変幻自在に変える神プロテウスにちなんでつけられたものである。
メタン生成菌は二酸化炭素と水素さえ手に入れば、自分が必要な有機物を合成できるが、問題は水素を手に入れることだった。真核生物が誕生したのはおよそ20億年前と考えられるが、それよりもかなり前には、酸素を放出するシアノバクテリアは進化していたので、20億年前にはすでに酸素のない環境は少なくなっていた。酸素があると水素はすぐに反応して水になってしまうのである。
従って、メタン生成菌にとっては、生きていく上で必要な水素を提供してくれるこの真正細菌は重要なパートナーとなり得るものだった。また、プロテオバクテリアにとっても自分の出した老廃物を除去してくれるメタン生成菌は、ありがたい存在だったと考えられる。この両者の関係は、図32-3bのようにメタン生成菌がこのプロテオバクテリアを細胞内に取り込むことによってより密接になった。
マーチンとミュラーは、このようにしてメタン生成菌の細胞内に共生するようになったプロテオバクテリアが、ミトコンドリアやヒドロゲノソームに進化したのではないかと考えたのだ。これが、「水素仮説」である。このような水素を通じた共生がきっかけになって、新しい型の生命が誕生した。
現在、メタン生成菌のような代謝を行なう真核生物はいないが、先に紹介したように、リヴェラとレイクによると、現在の真核生物の核ゲノムのなかで代謝に関係する「作業」遺伝子は、そのほとんどが真正細菌由来だという。従って、プロテオバクテリアが共生したあとで、真正細菌の遺伝子の多くが「作業」遺伝子として宿主のゲノムに取り込まれたものと考えられる。
マーチンとミュラーの「水素仮説」は、真核生物の起源を説明する可能なシナリオの一つに過ぎない。リン・マーグリスは、シアノバクテリアが放出した猛毒の酸素が充満して生物進化史上最初の大量絶滅を引き起こし、これに対処するためにミトコンドリアの共生が生まれたと考えた。しかし、そのような大量絶滅が起ったという証拠は見られない。
動物や植物など、日常われわれが見るほとんどの真核生物では、ミトコンドリアは酸素呼吸の場であり、このように酸素を消費してくれるミトコンドリアの共生は、猛毒の酸素に対する対抗措置として納得できそうに思われた。しかしながら、実際にはミトコンドリアは非常に多様である。
先にほとんどのヒドロゲノソームはミトコンドリアとは違って独自のゲノムをもたないと述べたが、実はニクトテラス・オヴァリスNyctotherus ovalisというゴキブリに寄生する繊毛虫のヒドロゲノソームはゲノムをもっており、それを調べてみると、ミトコンドリアと共通の祖先から進化したことが明らかになった。
「水素仮説」では古細菌のメタン生成菌と真正細菌のプロテオバクテリアが融合して真核細胞が誕生したと考えるが、水素はあくまでも共生関係が成立するきっかけになっただけで、その後の真核生物の代謝は主にプロテオバクテリアからもたらされた遺伝子によっているように思われる。
この仮説で想定しているメタン生成菌は、必ずしもこれまでに知られているメタン生成菌に限られるものではなく、水素分子に依存して自分で栄養を作り出せる独立栄養生物ということである。
真核生物の起源については、まだ不明なことが多いが、いずれにしてもおよそ20億年前に最初の真核生物が誕生した。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


