
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第21話
発生と進化
<表現型の可塑性>
文と写真 長谷川政美
◎ヘビの進化
ヘビはトカゲとともに爬虫類のなかの有鱗目に分類されるが、分子系統学からはトカゲのなかのオオトカゲ下目とイグアナ下目との共通祖先から進化したことが分かっている(図21-1)。 つまりヘビはトカゲのような四足動物から進化したのだ。
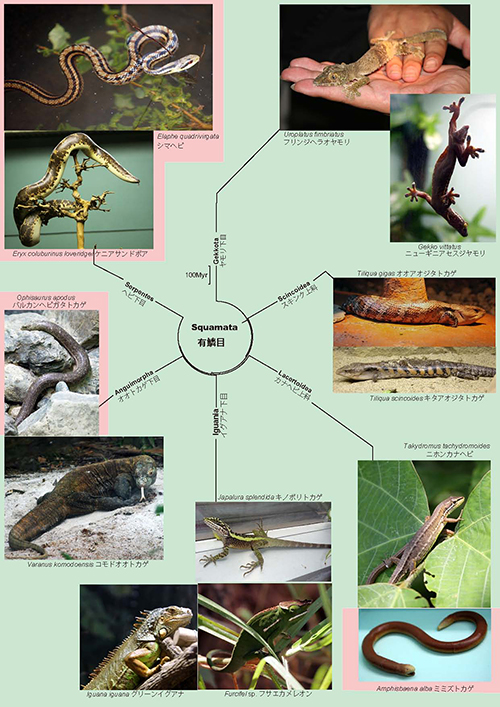
ヘビの祖先は大幅なからだの改造を行っている。4本の肢を失うだけでなく、からだを細長くして、それをくねらせることによって地上、樹上、水中などさまざまな環境で自由に移動できる独特の方法を獲得したのである。事実ヘビは爬虫類のなかで非常に繁栄しているグループであり、3,600種以上を擁している。
硬骨魚類から進化して陸上に進出した四足動物のなかには、いったん獲得した四肢を失うかたちで進化したものも多い。両生類のなかで四肢を失ったものが、無足目のアシナシイモリである(図21-2)。また、イモリと同じ有尾目のサイレンでは、四肢は残っているものの非常に短くなっている(図21-3)。さらに爬虫類のなかの有鱗目では、ヘビ以外のいくつかの系統でも4肢の退化が繰り返し起っている(図21-1のヘビガタトカゲやミミズトカゲなど)。


ヘビが進化するにあたって、4肢の退化とからだの伸長が同時に起っている。キリンの首が長くなる際には、頚椎の数は哺乳類一般の7個(ナマケモノなどでは違うこともある)のままなのに対して、ヘビでは200~400個もの脊椎骨、つまり椎骨をもつことによってからだを長くしているのだ(図21-4)。このような進化はホメオティック遺伝子の変異によって可能になっている。

「ヘッジホッグ遺伝子」という遺伝子は、最初ショウジョウバエの分節パターンを制御する遺伝子として同定された。この遺伝子の機能を失った変異体の胚には、ハリネズミの剛毛のような突起が密集するので、ヘッジホッグという名前がつけられた。
その後、この遺伝子ファミリーに属するいくつかの遺伝子が、ショウジョウバエだけでなく、脊椎動物でも見つかった。そのなかに、「ソニック・ヘッジホッグ遺伝子」と呼ばれるものがある。この遺伝子には、「エンハンサー」と呼ばれる遺伝子の発現を調節する領域があるが、ヘビのものには他の動物には見られない17塩基の欠失がある。エンハンサーは、魚類を含む広範囲の脊椎動物で保存されているが、ヘビで不活性化されているのだ。
アメリカのローレンス・バークレー国立研究所のエフゲニー・クヴォンらが、コブラのエンハンサー領域を切り取ってマウスの胚に導入したところ、肢のないマウスが生まれた。一方、ヒトやゼブラフィッシュのエンハンサー領域に組み換えたマウスでは、4肢はちゃんと生えたのだ。
彼らはさらに、ヘビで特異的に欠けている17塩基を追加したエンハンサー領域をマウスに導入したところ、今度は4肢の生えたマウスが生まれてきた。つまり、ヘビに肢が生えないのは、例のDNA領域で17塩基が欠損しているからだと考えられる。
ヘビに肢が生えない原因が、エンハンサーの不活性化だけだとすると、何らかの方法でこれを活性化することができれば、肢の生えたヘビが生まれるかもしれない。実験的に確かめられたわけではないが、どうもそのような可能性が高そうなのである。なぜならば、4肢をもったヘビの化石が発見され、当初それはヘビの祖先型であり、そのようなものから肢が退化して現在のヘビが進化したものと考えられた。ところが詳しく調べてみると、この肢をもったヘビは肢のないヘビから進化したもののようなのだ。従って、肢を失ったヘビで少なくとも1回は、4肢の復活が起っている。
◎ダーウィンフィンチの嘴
第3話でガラパゴスのダーウィンフィンチの嘴のかたちや大きさが、手に入る食べ物の違いで変化することを紹介した。近年のエボデボの進歩により、このような変化を引き起こす2つの遺伝的要因が明らかになってきた。この研究は、第6話に登場したピーター・グラントとローズマリー・グラント夫妻を含むアルハト・アブザーノフらのグループによって2006年発表された。
鳥類は嘴における細胞増殖因子としてBMP4というたんぱく質を使うことが知られているが、この遺伝子の発現量が嘴のかたちと強く相関しているのだ。
さらに、サボテンフィンチのように長く細長い嘴の鳥の胚は、太い嘴のものよりも、多くのカルモジュリンというカルシウム結合たんぱく質をもっていることが分かった。つまり、BMP4とカルモジュリンの発現量が自然選択の対象になっているということである(図21-5)。
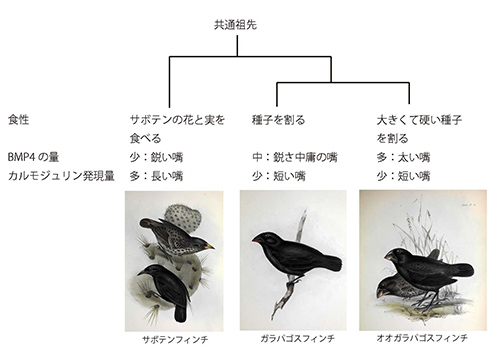
図21-5 ガラパゴスのダーウィンフィンチ類では、BMP4とカルモジュリンの発現量によって嘴のかたちと大きさが制御されている。フィンチの絵は、Gould (1841) http://darwin-online.org.uk/converted/pdf/1841_Birds_F9.3.pdfより。
◎同じ遺伝子の異なる表現型
日本にアカボシゴマダラというチョウがいる。このチョウはもともと奄美大島とその周辺の島々だけに分布していたが、1995年に埼玉県で目撃されて以来、関東から全国に急速に分布を拡大しつつある。これは奄美のものとは別亜種で、中国大陸から持ち込まれたものが広がったと考えられている。
2015年には、第12話と第17話にも登場した米澤隆弘君が、富士山の山頂付近でも目撃して報告している。
アカボシゴマダラの夏型成虫は、名前の通り赤い斑点をもっているが(図21-6a)、春型には赤い斑点はなく、翅全体が白っぽい(図21-6b)。


図21-6 アカボシゴマダラHestina assimilisの(a)夏型と(b)春型。遺伝的には同じチョウの夏型と春型の間で、環境条件や食草の違いなどでこのような違いが生み出される。
私も最初にこの春型個体を見たときには、アカボシゴマダラだとは思わず、あとで図鑑を調べてはじめて分かったことである。
同じようなことは、同じタテハチョウ科のアカマダラAraschnia levanaでも見られる。このチョウの春型の翅は黒い斑紋をもちオレンジ色であるが、夏型には白いバンドがあり、全体が黒色である。リンネはこの2つを別種としたほどである。
1875年にオーグスト・ワイズマンは、この違いが幼虫の発育中の温度の違いによるものであることを示した。その後の研究では温度だけでなく、日長も関係していることが明らかになっている。
私自身、アカボシゴマダラの春型と夏型の違いを生み出す機構がどの程度まで研究されているかを知らないが、このように遺伝的には同じものが環境や食べ物の違いで異なった表現型になるのは、しばしば見られることであり、表現型あるいは発生の可塑性と呼ばれる。表現型は、遺伝子型の単純な表現ではなく、遺伝子と環境の相互作用によって実現するものである。
北アメリカのNemoria arizonariaというシャクガ科のガは、春にカシの木で孵化すると、黄色でカシの花に紛れ込むすがたの幼虫になる(春型)。ところが同じカシの葉を食べるが花のない季節に生まれる夏の幼虫は、カシの枝に似ている(夏型)。
アメリカ・プリンストン大学のエリック・グリーンは、春型の幼虫はカシの若い葉を食べるために花に似るようになり、一方、硬い葉を食べる夏型は枝に似るようになることを明らかにした。遺伝的には同じはずの春型と夏型とが、食べ物の違いによって違った表現型を発現させているのである。
このような表現型の可塑性の顕著な例が、アリのカスト制であろう。女王アリと働きアリはどちらもメスであり、孵化したときには遺伝的に差がないが、その後の餌の量の違いで女王と働きアリに分かれるのである。
生物の多様性は、遺伝的な多様性だけでなく、環境の多様性によっても実現されている。進化とは、次世代に受け継がれるような形質の変化であるといわれる。それならば、表現型の可塑性は進化とは関係ないのであろうか。そんなことはない。なぜならば、ダーウィンとウォーレスがはじめて明らかにしたように、自然選択は表現型に対して働くからである。多様な表現型のレパートリーもまた自然選択の対象であり、環境に対する応答の仕方も進化してきたといえる。
◎タバコスズメガの実験
タバコスズメガManduca sexutaの幼虫は通常緑色であるが、黒色の変異体もいる。この黒色変異体の4齢幼虫に6時間42℃の熱ショックを与えると、5齢幼虫の体色は黒色から緑色までの幅広い表現型を示す。アメリカ・デューク大学のユウイチロウ・スズキとフレデリック・ナイハウトは、この段階での体色に対して13世代にわたって人為選択をかけてみた。熱ショックに対して緑色になった幼虫から出た成虫同士を掛け合わせるという選択を続けると、熱ショックを受けた幼虫はすべて緑色になった(緑色選抜系統)。逆に熱ショック反応のないもの同士を掛け合わせた系統は、温度に関わらず黒色のままだった(黒色選抜系統)。
黒色変異体では幼若ホルモンというホルモンの分泌量が野生型にくらべて少ないために黒化が起ると考えられるが、緑色選抜系統と黒色選抜系統では熱ショックによって分泌される幼若ホルモン濃度が選択の対象になったのだ。つまり、体色多型という可塑性の起源は幼若ホルモン濃度という量的なものであり、緑色の選抜を続けた系統では、熱ショックによる色の変化、つまり表現型の可塑性が増大する方向に進化が起り、逆に黒色選抜系統では可塑性が消失する方向に進化が起ったものと解釈できる。
スズキとナイハウトが行った「進化」は実験室でのものであったが、これに似たことは野外でも起こり得ると思われる。これは新たな突然変異なしで環境が誘導する変化だと考えられる。
スコット・ギルバートとデイビッド・イーペルによると、このように環境が誘導する変化は、突然変異遺伝子が集団に固定するというこれまでの一般的な進化様式にくらべて、進化的な新奇性の創出により効果的かもしれないという。突然変異の場合は1個体だけから出発するに過ぎないのだ。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


