
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第29話
生命の誕生
文と写真 長谷川政美
本連載の今回以降は、およそ46億年の地球の歴史のなかで、生物がどのように進化してきたかを見ていくことにする。生物の進化は、地球の進化と関連しながら進行してきたものであり、さらに生物進化が地球の進化にも影響を与えてきたことが分かるであろう。ただし、ここでは進化史上重要な事件を網羅するということではなく、私自身が進化生物学の研究を行なってきた過程で特に興味をもった問題を中心に紹介することを了承していただきたい。◎地球上の生命の起源
最初の生命が誕生した条件は、現在でも存在し、またこれまでもずっと存在していたのではないかとよく言われます。しかし、もし(とんでもなく大きな「もし」ですが)どこかの温かい水たまりにさまざまなアンモニアやリン酸塩が存在し、光、熱、電気などでたんぱく質が化学的に合成され、さらに複雑な変化が起きる準備ができたとしても、現在ではそのような物質はすぐに食べられたり、吸収されてしまうでしょう。でも、生命が誕生する前ならばそうではなかったはずです。
ダーウィンからジョセフ・フッカーに宛てた手紙 (1871)
ダーウィンからフッカーに宛てた上の手紙は、第1話で紹介したルイ・パスツールによる生物の自然発生説を否定した有名な実験からおよそ10年後のものである。ダーウィンは、現在では生命が自然に発生することはないが、地球上で1回だけ発生したはずだと考えたのである。
初期の地球上で生命が何回生まれたかということは、まだ答えが得られていない問題であるが、現在生きている生物はすべて一つの共通祖先から進化したことは、正しいであろう。なぜならば、あらゆる生物の遺伝的な仕組みが基本的に共通なので、一つの祖先から進化したとしか考えられないからである。しかし、最初に複数のタイプの生命が生まれたが、最終的に残ったのが一つだけだったのかもしれない。ダーウィンはいったん生命が生まれると、もはや別の生命は生まれにくくなると考えたのである。
地球はおよそ45億6700万年前に誕生した。最初の生命がいつ生まれたかは分からないが、40億年前~25億年前の始生代(表29-1)の間に生まれたことは確かである。
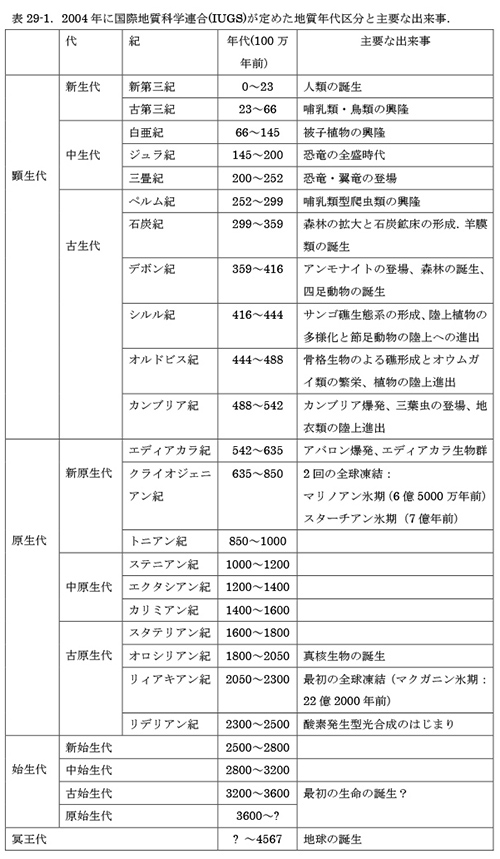
アメリカア・ラトガース大学のポール・フォーコウスキーによると、すでに38億年前にあとで出てくる酸素放出型とは違う型の光合成をする細菌がいた可能性を示唆するデータがあるという。生物のからだを構成する有機物は、すべて炭素を含む。炭素Cという元素には、原子量がそれぞれ12と13の12Cと13Cという2種類の安定同位元素(安定とは放射性同位元素のように別な元素に変わってしまわないという意味)が存在する。光合成により炭素を使って有機物を合成する際に、質量の小さな12Cのほうが13Cよりも取り込まれやすい(反応速度が速い)ので、この2つの同位元素の比を調べることによって、始生代における光合成細菌の痕跡を探すことができる。
ヨーロッパとアジアの間にある黒海の38億年前の黒鉛を調べると、12Cの同位体比が非生物的な自然界の平均値よりも高いので、この頃までに光合成細菌が進化していたことが示唆される。南アフリカとオーストラリア西部の35億年前の岩石からはもっとはっきりした生命の痕跡が見つかるという。
1920年代に、ロシアのアレクサンドル・イヴァノヴィッチ・オパーリン(図29-1)とイギリスのジョン・バードン・サンダーソン・ホールデンJ.B.S. Haldane(1892-1964)は、無酸素状態であった初期の地球上で形成された有機分子間の相互作用として生命が出現したという考えを提唱した。その後、生命の起源をめぐる研究は、実験科学としても大きな展開を続けてきた。

図29-1 アレクサンドル・イヴァノヴィッチ・オパーリンAleksandr Ivanovich Oparin(1894-1980)。1977年に京都で開かれた第5回・国際生命の起源学会で来日した際の写真。
最新の研究成果によると、海底から噴出するアルカリ熱水噴出孔というタイプの熱水噴出孔で最初の生命が生まれたという。このような熱水噴出孔はブラックスモーカーとも呼ばれ、第14話で出てきた、2つのプレートの境界域でマグマの噴出口が連なっている中央海嶺にある。大陸移動を引き起こす力はまた、最初の生命をも生み出したのだ。
噴出孔から出る熱水にはさまざまな鉱物が溶解しており、それをもとに有機物を合成する細菌が生まれた。細孔の迷路を通過する熱水の流れには、有機分子を濃縮する能力があるという(ニック・レーン著『生命、エネルギー、進化』斉藤隆央訳、みすず書房、2016 年)。
◎酸素発生型光合成細菌の出現
およそ25億年前の始生代から原生代に移行する時期に、二酸化炭素を変換して有機物を生み出す光合成を行なう生物が進化した。その新型生物は光合成の際に水分子を分解して酸素分子を放出したので、酸素発生型光合成と呼ばれる。それを行なったのがシアノバクテリアという細菌である。この細菌は、光のエネルギーを使った
6CO2 + 6H2O → C6H12O6 + 6O2
という反応を通じて二酸化炭素と水から炭水化物を合成し、酸素を放出する。シアノバクテリアが堆積して層状の構造をもったものをストロマトライトというが、それの古い化石は世界各地で見つかっている(図29-2)。

図29-2 ストロマトライトの化石(南アフリカ・マカパンスカットにて)。
シアノバクテリアがいつ出現したかという問題は、今でも議論が続いている。かつて西オーストラリアのおよそ35億年前の地層からフィラメント状の微化石が発見され、シアノバクテリアのものだと考えられていた。ところが、これは生物由来の構造ではないことが明らかにされた。シアノバクテリアがいつ出現したかという問題は、地球大気の歴史を考える上で重要であるが、およそ25億年前には出現したものと思われる。
シアノバクテリアにより大気中の二酸化炭素が消費され、逆に酸素が放出された。それまで地球上には酸素分子はほとんど存在しなかったが、それが突然増加したのだ。当時の海には海底火山や熱水噴出孔から噴き出した鉄分が多く溶けていた。シアノバクテリアが放出した酸素分子は海水中の鉄分と反応して酸化鉄に変わり、鉄鉱層となって海底に堆積した。世界各地で見つかる縞状鉄鋼層(図29-3)は、その頃にそのようにしてできたものである。酸素が海水中の鉄分と反応し尽くすと、海水中の酸素濃度がどんどん上昇した。

図29-3 オーストラリア北西部で見つかった25億年前の縞状鉄鋼層。シアノバクテリアの放出した酸素分子が海水中の鉄分と反応して酸化鉄に変わり、鉄さびが海底に堆積したもの。左に見えるのがくっついた磁石で、確かに鉄が多く含まれていることが分かる。このような鉄鉱石は世界各地で見られるもので、人類が鉄器を使った文明を発達させるのに貢献した(蒲郡市生命の海科学館・所蔵標本)。
それまでの嫌気的環境で育ってきた生物にとって、酸素はとんでもない猛毒であった。酸素は何とでもすぐに反応してしまうからである。そのためほとんどの生物が絶滅したという。これが細胞内共生説で有名なリン・マーグリス(図29-4)の考えた「大酸化事変」であり、猛毒の酸素による地球上最初の大量絶滅とされた。

図29-4 細胞内共生説の提唱者リン・マーグリスLynn Margulis (1938―2011)。1967年の論文は、当時天文学者のカール・セーガンCarl Sagan (1934―1996)と結婚していたので、リン・セーガンの名前で出版された。1997年、アメリカ・ウッズホールでの研究会にて。
葉緑体の細胞内共生の考えは、19世紀まで遡ることができる。1893年にドイツの植物学者アンドレアス・シンパーAndreas F.W. Schimper(1856-1901)が植物の葉緑体がシアノバクテリア(当時は藍藻と呼ばれていた)と似たようなかたちで分裂するのを観察して、葉緑体はシアノバクテリアが細胞内で暮らすようになったものではないか、と考えたのである。この考えは1905年に発表された、ロシアのコンスタンティン・メレシコフスキーKonstantin Mereschkowski(1855-1921)の「植物界における色素体(葉緑体のこと)の正体と起源について」という論文のなかで更に発展させられたが、マーグリスの論文が出るまでは広く受け入れられることはなかった。マーグリスはシンパーやメレシコフスキーの細胞内共生説を現代的なかたちで復活させたといえる。
マーグリスの説も当初は猛烈な反対に出会った。彼女は、論文を5年間にわたって主要な雑誌に15回も投稿し続けたが、ことごとく掲載を断られたという。彼女の50ページにもおよぶ長大な論文にはほとんど数理は出てこないが、最後にようやく理論生物学雑誌 Journal of Theoretical Biologyという数理的な論文の多い雑誌に掲載されたのである。細胞構造や系統樹の図にあふれた彼女の論文は、数理はなくても革新的な理論生物学であることが認められたのだ。その後、ミトコンドリアや葉緑体のもつDNAが、系統的には細菌のものと近縁であることが明らかになったこともあり、細胞内共生説は広く受け入れられるようになった。
マーグリスによれば、αプロテオバクテリアは猛毒の酸素を消費してくれるので、宿主はそれによって守られたというわけである。ところが、ニック・レーンによるとそうではないという。酸素を作り出す機構は光合成以外にもある。太陽から降り注ぐ紫外線は、生物なしでも光化学反応で水を分解して水素と酸素を作ることができる。水素は軽いので地球の引力から逃れて地球外に拡散してしまう(図29-5a)。
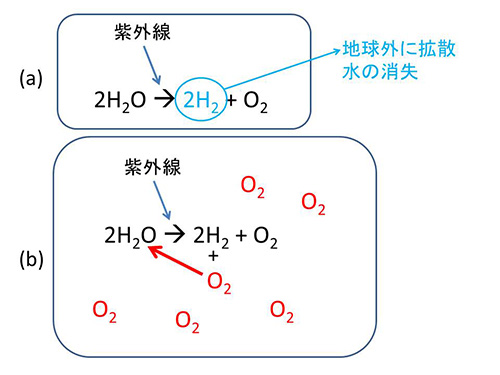
図29-5 紫外線による水H2Oの分解。分解されてできた水素は軽いためにすぐに地球外に拡散してしまったが(a)、シアノバクテリアによって酸素が作られるようになると、紫外線によって水が分解されても、できた水素分子はすぐに大気中の酸素分子(赤色)と反応して水に戻るようになった(b)。
シアノバクテリアが出現する前は、地球上でも同じようなことが起った。最初のうちは紫外線で作られた酸素は海水中の鉄分と反応したが、それが飽和する前に、シアノバクテリアによる酸素発生型光合成が進化した。レーンによると、このおかげで地球が火星や金星のような運命をたどらずに、水の惑星が守られたのだという。シアノバクテリアによって作られた酸素は鉄分と反応して酸化鉄を作ったが、そのような鉄分を使い果たしてしまうと、大気中の酸素濃度が次第に上昇した。酸素濃度がある程度高くなると、紫外線で水が分解されても、できた水素の大半は酸素と反応して再び水が作られるのである(図29-5b)。
紫外線により微量ではあるが環境中に酸素が作り出されていたので、それに対する防御策は酸素発生型光合成が現れる前にすでに進化していたはずだという。シアノバクテリアが酸素発生型光合成を始める前に、酸素に対する防御機構を進化させていなかったならば、自分の出した毒にやられて死滅してしまったであろう。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹



*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史 (BERET SCIENCE)
』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹
【バックナンバー】
第1話 「自然の階段」から「生命の樹」へ
第2話 リンネの階層分類
第3話 キュヴィエの新しい分類
第4話 共通祖先からの進化
第5話 偶然性の重視
第6話 自然選択の現場 ーガラパゴスフィンチ
第7話 なぜ多様な種が進化したか?
第8話 分子系統学の登場
第9話 ペンギンはなぜ北極にいないか
第10話 ウォーレスのマレー諸島探検
第11話 ペンギンの分布
第12話 ホッキョクグマの分布
第13話 ウェゲナーの大陸移動説
第14話 大陸移動説の拒絶と受容
第15話 大陸分断による種分化と
幸運に恵まれた移住
第16話 海を越えた移住
第17話 古顎類の進化
第18話 南極大陸を中心とした走鳥類の進化
第19話 進化発生生物学エボデボの登場
第20話 繰り返し要素の個性化と多様な形態の進化
第21話 表現型の可塑性
第22話 ジャンクDNA
第23話 少ない遺伝子
第24話 ヘモグロビンにおける調節
第25話 エピジェネティックス
第26話 獲得形質は遺伝するか?
第27話 美しいオス
第28話 性選択