
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第31話
古細菌と真核生物を結ぶ失われた鎖
文と写真 長谷川政美
◎真核生物の祖先とは?
古細菌が真核生物に近縁だということなので、古細菌から真核生物が進化したように見えるが、真核生物の大部分がもっているミトコンドリアという細胞内小器官(オルガネラともいう)は、マーグリスが主張したように「αプロテオバクテリア」という真正細菌が細胞内に共生して生まれたものである。また植物がもっている葉緑体という細胞内小器官は、「シアノバクテリア」という真正細菌が細胞内に共生して生まれたものである。従って、たいていの真核生物は実際には複数の生物の複合体だということになる。そうだとすると、αプロテオバクテリアが共生する以前の真核生物の祖先はどんなものだったのだろうか、という疑問がわく。それは古細菌と真核生物を結ぶ失われた鎖(ミッシングリンク)である。
トム・カヴァリエ・スミス(図31-1)は、1983年に、そのような失われた鎖の候補としてミトコンドリアをもたない一群の真核生物を挙げ、これらがミトコンドリア共生以前の祖先型真核生物ではないかと考えた。彼はこの仮想的な祖先生物を「アーケゾア Archezoa」と名付けた(図31-2)。

図31-1 アーケゾアの提唱者トム・カヴァリエ・スミスTom Cavalier-Smith (左:現オックスフォード大学教授)と筆者。1994年に筆者が彼の当時いたカナダ・バンクーバーを訪ねたときの写真。
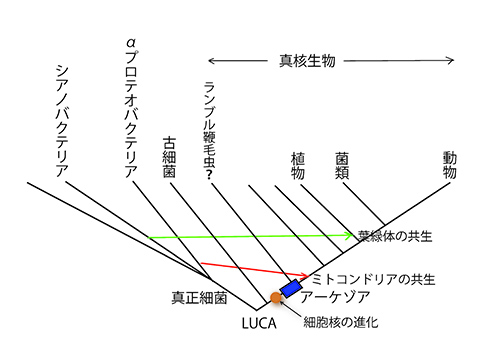
図31-2 真核生物の起源に関するアーケゾア仮説。カヴァリエ・スミスはミトコンドリアが共生する以前の祖先型真核生物をアーケゾアと呼んだ。
動物、植物、真菌類など真核生物の大部分は、細胞内にミトコンドリアをもつが、単細胞の原生生物のなかには、細胞核をもっていて真核生物であるにもかかわらず、ミトコンドリアをもたないものがいる。ヒトの腸内に寄生して下痢の原因になるランブル鞭毛虫(ディプロモナス類;図31-3)、ヒトの性感染症を引き起こすトリコモナス(パラバサリア類)、アユの寄生虫のグルゲアやヒトを含めた哺乳類に感染するエンケファリトゾーンなどの微胞子虫、ヒトに寄生してアメーバ性赤痢を引き起こすエントアメーバなどである。まずこれらの生物がアーケゾアの候補となった。
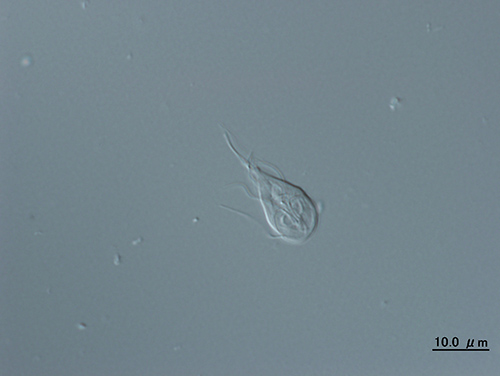
図31-3 ランブル鞭毛虫Giardia lamblia(©橋本哲男)。
◎探索の始まり
1990年代、私の研究室を含めて世界各地の研究室で、アーケゾアの探索が行われた。われわれのグループの中心は、当時私のいた統計数理研究所の助教授だった橋本哲男さんだった(図31-4)。

図31-4 ミクロス・ミュラーMiklós Müller(右)と橋本哲男さん(左後)。1998年にミュラー博士が統計数理研究所の筆者の研究室を訪ねてきた際の写真。
探索は、まずミトコンドリアをもたないいろいろな真核生物について、分子系統学的な解析から系統樹上でどのように位置づけられるかを調べた。その結果、エントアメーバはミトコンドリアをもったグループの内部に入ってしまうことがわかった。
第8話で出てきた図8-2の真核生物の系統樹マンダラで、アメーボゾアというグループに入るのだ。アメーボゾアの生物は、たいていミトコンドリアをもっている。従って、エントアメーバは、寄生生活に入ったあとでミトコンドリアを失ったものであると判断され、アーケゾアの候補から除外された。アーケゾアはミトコンドリアが共生する以前のものだから、図31-2に示すようにミトコンドリアをもったものよりも真核生物の系統樹の根元近くから分岐していなければならないのだ。
同じように、グルゲアなどの微胞子虫も調べられた。最初これは真核生物の系統樹の根元近くから分岐して、アーケゾアの有力な候補だと思われた。ところがこれは系統樹推定の間違いであることが明らかになった。微胞子虫も寄生性なので、自然選択上の制約が弱くなっているためなのか、非常に分子進化速度が速くなっていて、そのため間違った系統樹が得られていたのである。
現在では微胞子虫は真菌類の系統であり、寄生性になってからミトコンドリアを失ったものだと考えられている。こうして微胞子虫もアーケゾアの候補から除外された。
ランブル鞭毛虫(図31-3)とトリコモナスも当初、系統的な位置づけから、アーケゾアの候補にふさわしいと考えられた。真核生物のなかで最初にほかから分かれていたように思われたのだ。ほかにもミトコンドリアをもたないいくつかの真核生物が、系統樹の根元から分岐していて、アーケゾアの候補として残っていた。
◎アーケゾアが存在した証
アーケゾアの候補であることをはっきりと主張するためには、系統的な位置づけだけでなく、その生物がかつてミトコンドリアをもっていた痕跡が見られないことを示すことが必要である。この点をはっきりさせるために、橋本さんは1990年代の後半に、ニューヨークのロックフェラー大学のミコロス・ミュラー(図31-4)らの協力を得て、ランブル鞭毛虫やそのほかのミトコンドリアをもたない原生生物の遺伝子組成を調べた。
ミトコンドリアをもつ真核生物の核ゲノム中には、αプロテオバクテリアを共生体として受け入れた古細菌に由来する遺伝子のほかに、共生体がもっていた遺伝子の多くも保存されている。
ミトコンドリアは核ゲノムとは別に独自のゲノムをもっているが、それはαプロテオバクテリアのゲノムにくらべるとはるかに小さなものである。脊椎動物の場合、ミトコンドリアのゲノムは、22個のtRNA遺伝子、2個のリボソームRNA遺伝子、それに13個の呼吸鎖複合体サブユニット遺伝子しか含まないのだ。ところが共生する前のαプロテオバクテリアのゲノムは、これよりもはるかに多くの遺伝子をもっていたはずである。共生後にその多くは失われたが、残りはミトコンドリアから核に移行したのだ。従って、αプロテオバクテリア由来の遺伝子が核ゲノムのなかに見つかれば、その生物がたとえ現在ミトコンドリアをもっていなくても、かつてはこれをもっていた証拠になる。
そして、橋本さんらがランブル鞭毛虫やトリコモナスなどミトコンドリアをもたない原生生物の核ゲノムを調べたところ、αプロテオバクテリア由来の遺伝子が見つかった。
その頃、世界各地の研究室でも同じような研究が進んでいて、別のミトコンドリアをもたない原生生物でも次々とミトコンドリアの痕跡が見つかってきた。ミトコンドリアをもたない真核生物として知られている原生生物はほとんどが寄生性であるが、もともとミトコンドリアをもっていたが、寄生生活を送るようになってからこれを失ってしまったようなのだ。ほかの生物に頼って生きる寄生生活に入ると、必要なくなるものが出てくるのである。
◎仮説の否定
こうなると、アーケゾアというミトコンドリアが共生する前の祖先型真核生物が存在したという考えに疑念が生ずる。アーケゾアというミトコンドリアの共生を経験したことのない真核生物は地球のどこかに人知れずひっそりと生き延びているか、あるいはすでに絶滅してしまったという可能性はある。しかし、もう一つ別の可能性も考えられる。つまり、そもそもそのような生物は存在しなかったという可能性だ。ミトコンドリアの共生が、真核生物の誕生と密接に関わっているかもしれないということである。
アーケゾア仮説は、細胞核はもっているが、ミトコンドリアを共生させたことのない生物の存在を前提としている。しかしごく最近まで、細胞核の起源を合理的に説明する理論はなかった。もしかすると、ミトコンドリアの共生が細胞核を生み出すきっかけになったのかもしれない。そうであれば、アーケゾアという中間段階の生物は、もともと存在しなかったことになる。最近の研究の流れはそのような方向に向かいつつあることを次回以降で紹介しよう。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


