
870万種ともいわれる地球上の多様な生き物たち。
まだ私たちはそのごく一部しか知らないが、
実に多くのことが明らかにされてきてもいる。
進化生物学者である著者が、
世界中で長年撮りためた貴重な写真と文章で
思いのままに「生き物」を語る。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

進化の目で見る生き物たち
第18話
退化と中立進化
文と写真 長谷川政美
進化の過程で、特定の器官が縮小したりなくなったりすることを「退化」という。また適応度には差がないような形質に置き換わることを「中立進化」という。今回は退化と中立進化を取り上げてみよう。
◎「進化」と「退化」
「進化」という日本語は英語の「Evolution」の訳として、明治時代初期に作られた。Evolutionには「展開」という意味があり、それを「進化」と訳したわけであるが、そこには「進歩」という考えが入っているように思われる。確かに西洋の進化論者のなかには、高みを目指す進化という考えがあった。古代ギリシャのアリストテレスの「自然の階段」という捉え方である。アリストテレスは進化を考えたわけではないが、近代の進化学者は「下等な」生物から「高等な」生物へと階段を上るように進化すると考えたのである。ところが、ダーウィンの考えた「進化」はそれとは真っ向から対立するものであった。
ダーウィンの考えでは、ヒトはチンパンジーと共通の祖先から進化したが、チンパンジーからヒトが進化したのではない。彼の考えでは、共通祖先からヒトが進化したように、チンパンジーも進化したのである。ただ、2つの系統で進化の方向が違っていただけなのだ。
ダーウィン自身は1859年に出版した『種の起原』では、Evolutionという言葉を使わず、これを「変化をともなう由来(Descent with modification)」と表現している。彼は1872年に出版した『種の起原』第6版ではじめてEvolutionを使っているが、それでも本全体で10回しか使っておらず、Descent with modificationという表現が相変わらず多い。しかし、この言葉は「進化」の代わりに使うには長過ぎるので、一般にはEvolutionがよく使われるようになったのである。
ダーウィンは『種の起原』(1)のなかで、大西洋のマディラ島の甲虫の翅が退化して飛べなくなったことを取り上げている。この島に生息する550種の甲虫のうち220種は翅に欠陥があって飛ぶことができない。特にこの島固有の29属では、そのうちの23属ではすべての種が飛ぶことができない。ダーウィンによると、風の強いこの島では、よほど強力な飛翔力をもたない限り海に吹き飛ばされてしまうので、むしろ翅を退化させた方が有利だったのではないか、という。
このように、退化にも積極的な意味があり、「進化」と「退化」は対立する概念ではなく、退化も進化の一つの形態と捉えるべきである。生き物が使うことのできる資源には限りがあるので、一つの機能を強化するためには、別の機能を犠牲にしなければならない。例えば、チーターは陸上の肉食獣のなかで最速のランナーであるが、その分、頭部や顎の力が弱くせっかくの獲物をライオンやハイエナなどに横取りされることがある。どのような機能が強化され、逆にどのような機能が退化するかは、その生き物が置かれた生態的な状況によって決まる。
◎機能転換
ある器官が小さくなって、一見退化とまぎらわしいものもある。ハエやカなど双翅目昆虫では、後翅が小さくなって「平均棍」と呼ばれるものになっている(図18-1a、b)。



ところが、甲虫などの鞘翅目では、飛翔にはもっぱら膜状の後翅が使われ(図18-3)、硬い鞘状の前翅は飛んでいないときには柔らかい後翅を保護するとともに、飛ぶときにはからだを流線型にする役割もある。

◎オサムシにおける後翅の退化
双翅目昆虫で後翅が小さくなっているのは退化ではなく、機能転換であることを見た。しかし、オサムシ亜科の多くでは実際に後翅が退化している。オサムシ亜科は全ての甲虫が属する鞘翅目に分類されるが、鞘翅目では図18-3で示したように、前翅は硬い鞘になっていて、飛ぶためには使われない。従って、後翅が退化するということは、飛翔能力を失うことを意味する。
オサムシの系統進化を長年研究してきた大澤省三(元・JT生命誌研究館)(2)のグループは、オサムシ亜科の分子系統樹から、この分類群における後翅退化のシナリオを次のように描いている(3)。 図18-4にオサムシ亜科の分子系統樹を示した。
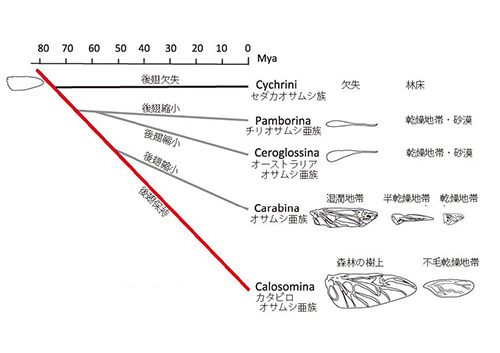
一方、カタビロオサムシ亜族ではおよそ半分の種では後翅が保持されていて、飛翔能力がある。後翅が保持されているのは、森林の樹上に生息するものであって、樹上の毛虫などを捕食するが、乾燥地帯に生息するものはたいてい後翅が小さく飛べなくなっている。
オサムシ亜科の共通祖先から出発してセダカオサムシ族と分かれたあとのカタビロオサムシ亜族に至る系統(赤線で示した)から、チリオサムシ亜族+オーストラリアオサムシ亜族、オサムシ亜族が順次枝分かれしている。
チリオサムシ亜族とオーストラリアオサムシ亜族は、それぞれ南アメリカとオーストラリアの乾燥地帯に生息し、後翅が縮小している。また、オサムシ亜族では、湿潤地帯に生息するものの後翅は比較的大きく、乾燥地帯のものでは退化が進んでいる。
以上のことから、オサムシ亜科における後翅の退化は、系統的に決まったグループだけで起こっているのではなく、生態的な条件に従って繰り返し起こってきたことが分かる。
オサムシ亜科はゴミムシ亜科から枝分かれしたが、この2つの亜科の共通祖先は完全な後翅をもっていた。ところがそこからカタビロオサムシ亜族に至る系統から順次分岐していった系統で、生息する環境にあわせて後翅を退化させたものが繰り返し現れたのである。
図18-5にオサムシ亜族の3種の甲虫について、湿地と乾燥地帯で後翅の大きさが顕著に異なることを示した。

それは、乾燥地帯では樹木が少ないので、空を飛ぶメリットが少なく、地上で小動物を捕食するようになったために、後翅の退化が進んだと考えられる。
飛翔するためには後翅とそれを動かす飛翔筋が必要であるが、それを維持するためにはコストがかかる。必要のないコストを省くという退化も、一種の適応進化といえる。
ただし、オサムシ亜族内では湿潤地帯のものは後翅が一見完全のように見えてもすでに飛べなくなっているという。従って、単に湿度が後翅の縮小速度を左右しているだけなのかもしれない。
◎中立進化
自然選択の対象となる「適応度」という尺度では違いがない形質に置き換わる進化を「中立進化」という。このことは、木村資生(1924~1994)が1968年に提唱したように分子レベルの進化で特に顕著に見られる(4)。分子レベルでは、適応度に差のない中立的な変異がたまたま集団に広まることによって起こる進化が多いというのである。分子レベルでも適応的な進化は見られるが、木村の分子進化の中立説は、分子レベルの進化を捉える基本的な枠組みとして次第に定着した。
実はダーウィンも中立的な進化があることには気がついていた。彼は、機能を失った痕跡器官は種の由来を明らかにする手掛かりとして有効だと述べている。分子系統学で用いられるDNAにも、中立的な進化が多いということで似たような性質があるのだ。
表18-1は遺伝コード表である。
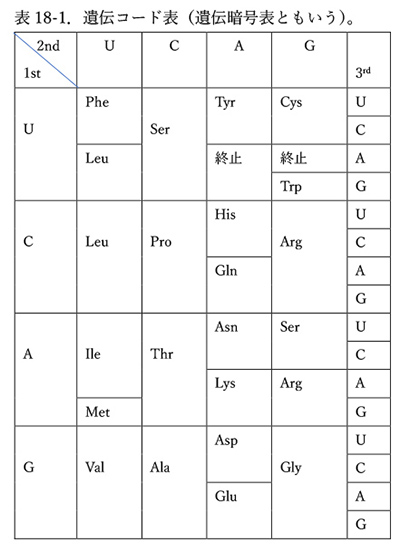
このコード表は1960年代に確立されたが、ヒトから細菌に至るまであらゆる生物でほとんど共通だということが分かってきたため、「普遍コード表」といわれる。ここで「ほとんど」と書いたのは、詳しく見るといくつかの生物では普遍コード表と少しだけ違ったコード表が使われていることも分かってきたからである。違っているといっても、普遍コード表が少し改変されただけであることは明らかなので、コード表の変異は、むしろあらゆる生物が一つの共通祖先から進化したという根拠にもなっている(5)。
真正細菌の一種にマイコプラズマ(Mycoplasma)がある。マイコプラズマのゲノムは55万~140万塩基対程度しかなく、大腸菌の420万~470万塩基対にくらべると小さい。マイコプラズマでは、普遍コード表の終止コドンUGAがトリプトファン(Trp)を指定するコドンに変わっている。
遺伝コード表のこの変化が、中立的な進化であるという議論がある(6)。
マイコプラズマのゲノムでは、G、C → A、T方向への突然変異率が逆方向への突然変異率よりも高くなっていて、同じアミノ酸を指定するのであれば、G、CよりもA、Uを多く含むコドンが使われる傾向がある。
普遍コード表の終止コドンとしては、UAA、UAG、UGAの3種類があるが、突然変異率の偏りの結果、ゲノム上ではTGA → TAAという変異が重なって、UGAが終止コドンとしては使われなくなったことが考えられる。そのような状況で、UGGを認識してトリプトファンを取り込むTrp-tRNA遺伝子が重複し、そのうちの一方の遺伝子がUGAコドンを認識するように進化した結果、UGAがトリプトファンを指定するコドンになったと考えられる(7)。
このようなコード表の変化の途中経過はすべて中立的な変異ととらえることができる。分子進化の中立説が提唱された当時、中立的な変異は生物学的な進化とは関係ないので、木村が見つけたことはたとえ正しいとしても生物学的には意味がない、という議論があった。しかし、退化が進化の一形態であるのと同様に、中立進化も進化の一つの形態であり、このような進化様式の存在は、生物学的にも大きな意味をもつ。
遺伝コード表の変化は、それによって新しい地平を切り開くような革新的な進化に結びつくものではないかもしれないが、中立的な変異があることによって、進化の可能性が広がることは確かであろう。図18-6はそのことを象徴的に表わしている。

このように多様な中立変異があると、さまざまな方向へ進化できる可能性が高まるのだ。もしかすると、多くの革新的な進化は中立進化を介して起こるのではなかろうか。
【引用文献】
1. チャールズ・ダーウィン(1859)『種の起原』(八杉竜一訳、岩波書店、1963年)
2. Osawa, S., Su, Z.-H., Imura, Y. (2004) “Molecular Phylogeny and Evolution of Carabid Ground Beetles”, Springer.
3. Imura, Y., Tominaga, O., Su, Z.-H., et al. (2018) Evolutionary history of carabid ground beetles with special reference to morphological variations of the hind-wings. Proc. Jpn. Acad. B94, 360-371.
4. 木村資生(1986)『分子進化の中立説』(向井輝美・日下部真一訳、紀伊國屋書店)
5. Osawa, S. (1995) “Evolution of the Genetic Code”, Oxford Univ. Press.
6. 木村資生(1988)『生物進化を考える』岩波書店.
7. Yamao, F., Iwagami, S., Azumi, Y., et al. (1988) Evolutionary dynamics of tryptophan tRNA in Mycoplasma capricolum. Mol. Gen. Genet. 212, 364-369.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




