
870万種ともいわれる地球上の多様な生き物たち。
まだ私たちはそのごく一部しか知らないが、
実に多くのことが明らかにされてきてもいる。
進化生物学者である著者が、
世界中で長年撮りためた貴重な写真と文章で
思いのままに「生き物」を語る。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

進化の目で見る生き物たち
第6話
クマ科の進化
文と写真 長谷川政美
前回はヒグマの進化についてお話しした。今回は話題をもっと広げてクマ科全体の進化を取り上げることにする。その中で、絶滅した北アメリカとヨーロッパの巨大なクマや本州に生息するニホンツキノワグマの起源についても論ずる。◎クマ科全体の系統樹
現生のクマ科は3つの亜科に分類される。ジャイアントパンダ亜科、メガネグマ亜科、クマ亜科である。図6-1はミトコンドリアDNAデータから推定されたクマ科全体の系統樹マンダラである。
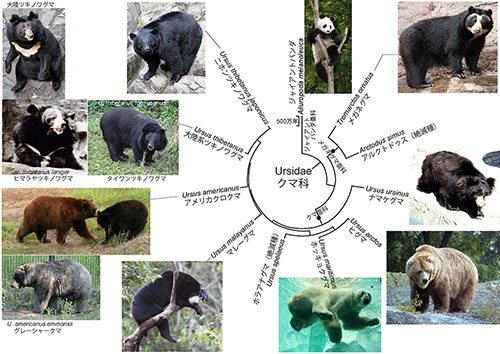
◎メガネグマ亜科の絶滅した巨大なクマ
メガネグマ亜科の現生種は南アメリカのメガネグマ一種だけだが、絶滅種としては北アメリカにアルクトドゥスがいた(図6-2)。

図6-1の中のアルクトドゥスの位置づけは化石の古代DNA解析によるものである(2)。ただし古代DNA解析とはいっても、化石そのもののDNAを調べるのではなく、絶滅した動物が生きていた環境に遺したDNAを調べる、いわば「古代環境DNA解析」と呼ばれる手法も最近開発されてきた(3)。化石のDNAを調べるには通常は化石に穴をあけてその中のDNAを取り出すが、環境DNA解析ならば貴重な化石を傷つけずにすむのだ。
メキシコの複数の洞窟の更新世後期(1万6000年前~1万4000年前)の地層から採取された環境DNAとしてアルクトドゥスとアメリカクロクマのDNAが見つかった。これらをカナダで見つかった2万2000年前~5万年前のアルクトドゥスの化石から採取されたDNAや現生のアメリカクロクマのDNAと比較したところ、メキシコ更新世後期のアメリカクロクマは現生のアメリカクロクマに近縁であるが、2地点のアルクトドゥスは遺伝的に互いにかなり離れていることが分かった。アルクトドゥスには多様な系統があったのである。
なぜアルクトドゥスは絶滅したのだろうか。彼らは最終氷期が終わったおよそ1万1000年前に絶滅したが、同じ頃に地上性のオオナマケモノをはじめとする北アメリカの大型哺乳類の多くも絶滅した。この絶滅には気候変動による環境変化が関わっている可能性があるが、アメリカ先住民の進出が関わっている可能性もある。
◎ツキノワグマの仲間
ツキノワグマのグループには、マレーグマ、アメリカクロクマ、ツキノワグマが含まれる。アメリカクロクマは北アメリカに分布するが、「クロクマ」という名前にもかかわらず、体色は黒色以外に、図6-1のアラスカのグレーシャークマのような青色からシナモン色までさまざまである。
ツキノワグマは、別名アジアクロクマとも呼ばれるように、西はイラン、アフガニスタンから東は日本まで、アジア全域に広く分布する。前回、北海道のヒグマは3つの系統に分けられ、それぞれが世界のヒグマの中の別々の系統に由来すること、またかつて本州にいた巨大なヒグマは、北アメリカのグリズリーに近縁な道南ヒグマがおよそ14万年前の氷河期に津軽海峡を渡って渡来したものであることを紹介した。
一方、分子系統学的解析から本州と四国に分布するニホンツキノワグマ(九州にも分布していたが絶滅したとされている)は、非常に古い起源をもっていることが明らかになった(1, 4)。ニホンツキノワグマはツキノワグマ全体の系統樹の中で最初にほかから分かれたのである。ニホンツキノワグマに対して、台湾ツキノワグマを含む大陸系のツキノワグマは、系統的にまとまった一つのグループを形成しているから、ニホンツキノワグマはツキノワグマ全体の中でも独自の系統なのだ(図6-1)。
ツキノワグマ進化の初期には、この種はユーラシア大陸全体に広く分布しており、日本にも分布していたと考えられる。ツキノワグマがアメリカクロクマと分かれて独自の進化の道を歩み始めたのはおよそ390万年前、ニホンツキノワグマが大陸のツキノワグマと分かれたのがおよそ150万年前の更新世(260~1.2万年前)だったと推定される。更新世は地球の寒冷化が進んだ時代であり、その間にツキノワグマの生息地である森林が次第に縮小し、多くの地域集団が絶滅したと考えられる。ニホンツキノワグマはこの時代を生き抜いてきた集団だったのだ。
ただし、ニホンツキノワグマに関しては不可解な問題がある。ツキノワグマの中で最初にほかから分かれた古い系統なのに、グループ内の多様性が意外に低いのである。特定のグループのDNAの多様性から、最後の共通祖先が生きていた年代(tMRCA: Time of the most recent common ancestor)が推定できる。大陸のものを含めたツキノワグマ全体のtMRCAにくらべて、ニホンツキノワグマのtMRCAは10分の1に過ぎないのである。このことは、ニホンツキノワグマは日本に渡来して以来ずっと大きな個体数を保持し続けたのではなく、最近になって個体数を爆発的に増やしたことを示唆する(1)。
これには、前回紹介した本州ヒグマの存在が影響していたと思われる。本州ヒグマの一つの系統はおよそ14万年前に北海道から本州に渡来したと考えられるが、34万年前にはすでに別の系統がいた。このヒグマがニホンツキノワグマを圧迫していた可能性があるのだ。更新世が終わる1万2000年前頃に本州のヒグマが絶滅したのを機に、ニホンツキノワグマ繁栄の時代を迎えたのかもしれない。
◎ホラアナグマ
前回ヒグマとホッキョクグマが非常に近い親戚であることをお話ししたが、この2種の姉妹群が絶滅したホラアナグマである。ホラアナグマは更新世後期のヨーロッパとアジア西部に生息していたが、およそ2万4000年前に絶滅した。このクマもアルクトドゥス同様に巨大だった(図6-3)。

一方同じ頃ヨーロッパにはヒグマも生息していたが、こちらは5万年前から生息数はあまり変わっていない。ホラアナグマとヒグマの間のこの違いは何によるのだろうか。今から5万年前頃からヨーロッパ各地に現生人類(Homo sapiens)が拡がっていたので、これがホラアナグマ絶滅と関係あるのではないかと考えられるが、それならばなぜヒグマも同じように減らなかったのかという謎が残る。
ホラアナグマもヒグマと同様に冬眠をしていた。スペインのホラアナグマが代々冬眠に使っていた洞窟には、ときには数千個体分の遺骨が蓄積していることがある。これらの骨に残っているDNAの解析からさまざまなことが明らかになってきた(7)。これらのホラアナグマのDNAからは、同じ血縁のグループが何世代にもわたって同じ洞窟を使っていたことが分かる。同じ血縁のオスもメスも自分が生まれた洞窟に戻って冬眠していたと見られるのだ。一方、ヒグマが冬眠に使っていた洞穴についても同様のことが調べられたが、そのようなことは認められず、ヒグマの場合、冬眠に使う洞窟の選び方はホラアナグマにくらべると柔軟であった。
特定の洞窟に対するこだわりの有無が、現生人類の拡散によって一方が絶滅し、他方が生き残った原因だった可能性があるのだ。初期の現生人類は洞窟に住むことが多く、洞窟をめぐってホラアナグマはヒトと競うことになった。更に特定の季節に特定の洞窟に帰巣するホラアナグマの習性は、現生人類による狩りを容易にしたのではなかろうか。
◎ヒグマのゲノムに遺されたホラアナグマのゲノム
ホラアナグマはおよそ2万4000年前に絶滅したが、そのゲノムの一部はヒグマのゲノムの中に今でも遺っている。ゲノムが解読された結果、ヒグマのゲノムの0.9~2.4%がホラアナグマに由来するものであることが分かったのである(8)。
種はその中だけで生殖が行われる集団と定義されるが、前回も見たように実際には種の壁を越えた交雑はしばしば見られる。ヒグマのゲノムの数%がホラアナグマに由来するということは、現生人類(Homo sapiens)とネアンデルタール人(Homo neanderthalensis)の関係に似ている。アフリカで進化した現生人類がユーラシアに進出した際にネアンデルタール人に出会い交雑した。ネアンデルタール人はその後絶滅したが、そのゲノムの一部はわれわれのゲノムにも残っているのである(9, 10)。
【引用文献】
1. Wu, J., Kohno, N., Mano, S., et al. (2015) Phylogeographic and demographic analysis of the Asian black bear (Ursus thibetanus) based on mitochondrial DNA. PLoS ONE 10(9), e0136398.
2. Krause, J., Unger, T., Noçon, A., et al. (2008) Mitochondrial genomes reveal an explosive radiation of extinct and extant bears near the Miocene-Pliocene boundary. BMC Evol. Biol. 8, 220.
3. Pedersen, M.W., De Sanctis, B., Saremi, N.F., et al. (2021) Environmental genomics of Late Pleistocene black bears and giant short-faced bears. Curr. Biol. 31, 2728-736.
4. Bit, A., Thakur, M., Singh, S.K., et al. (2021) Assembling mitogenome of Himalayan black bear (U. t. laniger) from low depth reads and its application in drawing phylogenetic inferences. Sci. Rep. 11, 730.
5. Stiller, M., Baryshnikov, G., Bocherens, H., et al. (2010) Withering away—25,000 years of genetic decline preceded cave bear extinction. Mol. Biol. Evol. 27, 975-978.
6. Gretzinger, J., Molak, M., Reiter, E., et al. (2019) Large-scale mitogenomic analysis of the phylogeography of the Late Pleistocene cave bear. Sci. Rep. 9, 10700.
7. Fortes, G.G., Grandal-d’Anglade, A., Kolbe, B., et al. (2016) Ancient DNA reveals differences in behaviour and sociality between brown bears and extinct cave bears. Mol. Ecol. 25, 4907-4918.
8. Barlow, A., Cahill, J.A., Hartmann, S., et al. (2018) Partial genomic survival of cave bears in living brown bears. Nature Ecol. Evol. 2, 1563-1570.
9. スヴァンテ・ペーボ (2015) 『ネアンデルタール人は私たちと交配した』 野中香方子訳、文藝春秋社.
10. 長谷川政美(2020)COVID-19とネアンデルタール人の遺伝子.『ウイルスという存在』第3話.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。





{kind=link}
{kind=link}