
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第27話
泡沫状ウイルスと
宿主哺乳類の共進化
文と写真 長谷川政美
この連載でこれまで、ウイルスは宿主にくらべて進化速度が速いので、地質学的に古い年代まで遡って進化の歴史を追うことは難しいが、内在性ウイルス様配列は宿主と同じような速度で進化するので、ウイルス進化の歴史を追うのに使うことができることを紹介した。今回は、内在化していない外因性ウイルスであっても、およそ1億年の進化の歴史を遡るのに使うことができる例を紹介しよう。
◎泡沫状ウイルスと宿主の共進化
泡沫状ウイルス(foamy virus)というウイルスがある。ヒトT細胞白血病ウイルスやヒト免疫不全ウイルス(HIV)と同じくレトロウイルス科に属するが、ほかのレトロウイルスとは別の独自のスプマウイルス亜科(Spumavirinae)に分類される。感染した細胞を培養すると泡状に細胞変成することからつけられた名前であるが、“spuma”とはラテン語で「泡」という意味である。ちなみに泡沫状ウイルス以外のレトロウイルスは、オルトレトロウイルス亜科(Orthoretrovirinae)に分類される。ここで“ortho”とはギリシャ語で「正しい」あるいは「真正」という意味であり、泡沫状ウイルスがレトロウイルスとしては変わっていることを示している。
このウイルスはさまざまな動物から採取されるが、感染による病理的な症状は知られていないので、最近まであまり注目されないウイルスだったが、進化学的に面白い話題を提供する。
泡沫状ウイルスには宿主ゲノムに組み込まれて遺伝する内在化が知られている例は少ないが、外因性ウイルスを使っても1億年近い進化の歴史を遡ることができるのだ。
哺乳類でも内在性泡沫状ウイルス配列はあまり知られておらず、これまでアイアイとフタツユビナマケモノでしか見つかっていない。
泡沫状ウイルスのゲノムにはPol(ポリメラーゼ)遺伝子があり、その中に逆転写酵素などいくつかの酵素がコードされている。イギリス・オックスフォード大学のアリス・カツオウラキスらのグループは、図27-1で示したように、逆転写酵素のアミノ酸配列(162アミノ酸)を使ってウイルスの系統樹(赤色)と宿主哺乳類の系統樹(黒色)を描き、二つの系統樹をくらべた。
その結果、ウイルスと宿主の系統樹が驚くほどよく一致していることが分かった(1)。このことは、宿主の種分化にあわせてウイルスも一緒に種分化してきたことを示す。このことを共進化(coevolution)あるいは共種分化(cospeciation)という。
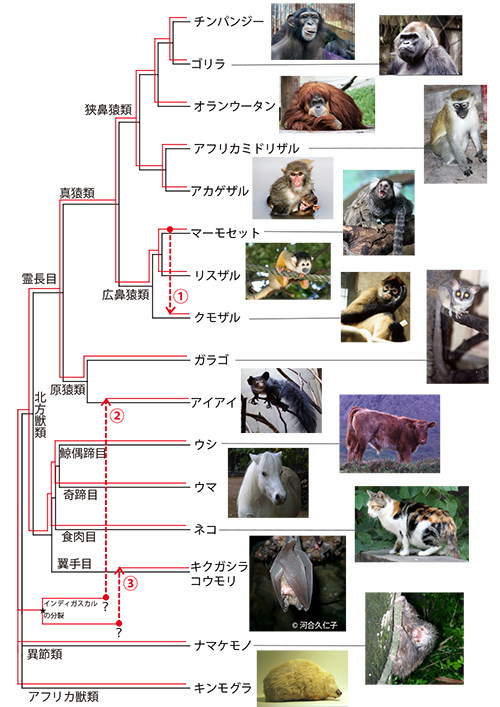
ただし、以下の3点において両者の枝分かれの順番に食い違いが見られる。1つは広鼻猿類(新世界ザル)の中である。マーモセットとリスザルはクモザルにくらべてお互いに近縁な関係にあるが、マーモセットのウイルスがリスザルよりもクモザルのものに近縁なのである。
このことは矢印①で示すように、マーモセットの祖先と共生していた泡沫状ウイルスが、宿主を換えてクモザルの祖先と共生するようになったことを意味する。逆に、クモザルのウイルスがマーモセットに移ったのかもしれないが。真獣類の進化の歴史を通じて、基本的には泡沫状ウイルスは宿主哺乳類と共進化してきたが、まれに宿主を乗り換えることもあったのだ。
2つの系統樹間で見られる残り二つの食い違いは、マダガスカルのアイアイと中国のキクガシラコウモリである。この二種と共生するウイルスは、系統樹上で宿主が位置する場所ではなく、お互いに一つのグループとして独自の系統を作っている。
アイアイは霊長目、キクガシラコウモリは翼手目であり、系統的に離れた動物であるにもかかわらず、お互いに近縁なウイルスが共生しているのである。しかも、霊長目と翼手目は真獣類の中で北方獣類という大きなグループに属するが、この二種類の動物と共生するウイルスは、ほかの北方獣類と共生するウイルスとは系統的には離れたものである(図27-2)。
図27-1で示した泡沫状ウイルスの系統樹が正しいとすると、次のようなシナリオが考えられる。
ウイルスが採取されたアイアイはマダガスカル、キクガシラコウモリは中国に棲息するものである。この系統的に離れた二種類の動物に近縁なウイルスが共生するためには、祖先が同じ環境で暮らしていたということが必要である。現在は遠く離れたマダガスカルと中国を結びつけるものがあり得るのであろうか。ここでカツオウラキスら大胆な仮説を提唱している。
彼女らは、マダガスカルと中国を結びつけるものは大陸移動だという。今から1億3000万年以上前には、現在のインドを含めた南半球の大陸はゴンドワナ超大陸として一つにまとまっていた。それが1億3000万年前になると、インドとマダガスカルをあわせた陸塊(インディガスカルという)がそれまで陸続きだった現在のアフリカ、南極、オーストラリアから分かれた。
その後1億500万年前にはアフリカと南アメリカが分離し、大西洋が生まれた。インディガスカル陸塊はおよそ8500万年前にインドとマダガスカルとに分離し、その後マダガスカルはほとんどそのままの位置にとどまったが、一方のインドは北上し、およそ4500万年前にユーラシアと陸続きになった(2)。
カツオウラキスらは、インディガスカル陸塊に棲息していた現在は絶滅した未知の動物に泡沫状ウイルスが共生していて、それがアイアイの祖先とキクガシラコウモリの祖先に宿主替えしたのではないかという(図27-3)。
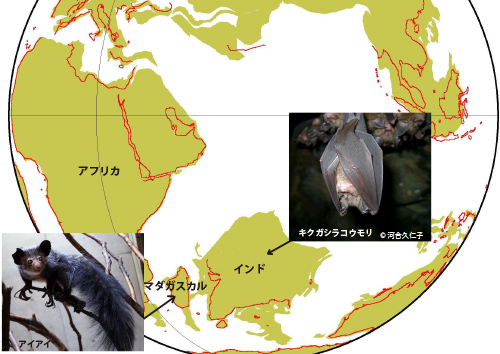
インディガスカル陸塊がマダガスカルとインドに分裂したおよそ8500万年前には、そこにはまだアイアイの祖先もキクガシラコウモリの祖先もいなかった。アイアイつまりキツネザルの祖先は、今からおよそ6000万年前にアフリカから海を渡ってやってきたと考えられているのである(2)。
カツオウラキスらによると、インディガスカル陸塊が分離した後で、マダガスカルとインドにいた未知の哺乳類から、アイアイの祖先とキクガシラコウモリの祖先に泡沫状ウイルスが引き渡されたものであろうという(図27-1の矢印②と③)。
このようにインディガスカルの分裂が動物の種分化のきっかけになったという例はカエルなどでは知られているが(2)、哺乳類では知られていない。
この仮説は壮大で魅力的なものであるが、分子系統樹推定は必ずしもいつも正しい系統樹を与えるとは限らない。例えば特定の系統でウイルスがほかとは違った選択圧を受けるなどした場合に、間違った系統樹が得られることがある。
この研究で用いられたアイアイとナマケモノの泡沫状ウイルスの配列は内在性ウイルスのものであり、内在化したことによって違った選択圧を受けてきた可能性がある。この仮説が正しいかどうかを確かめるためには、さらに多くの種、とりわけアジアのさまざまなコウモリやマダガスカルのさまざまなキツネザル、と共生する泡沫状ウイルスの詳しい解析が望まれる。
◎時間スケールを変えると
進化速度が変わる
第7話で短い時間スケールで観測した進化速度(ゲノムの塩基置換速度)が、長い時間で観測したものよりも高くなることを見た。それには時間が長くなると多重置換が重なって見かけ上進化速度が低くなることと、短い時間では弱有害の変異が残っているが、時間が経つと負の自然選択でそれらが次第に除かれていくという2つの原因が考えられた。
図27-1の泡沫状ウイルスゲノムの系統樹(赤色)は、宿主進化の時間スケールにあわせて描いたもので、ウイルスの塩基置換数を反映したものではない。図27-4であらためて泡沫状ウイルスの系統樹の枝の長さを塩基置換数に比例させて描いた(赤色)。それを宿主の分岐年代に比例させた黒色の系統樹と対比すると、泡沫状ウイルスの進化速度の時間依存性に関して興味深い様相が明らかになる(3)。
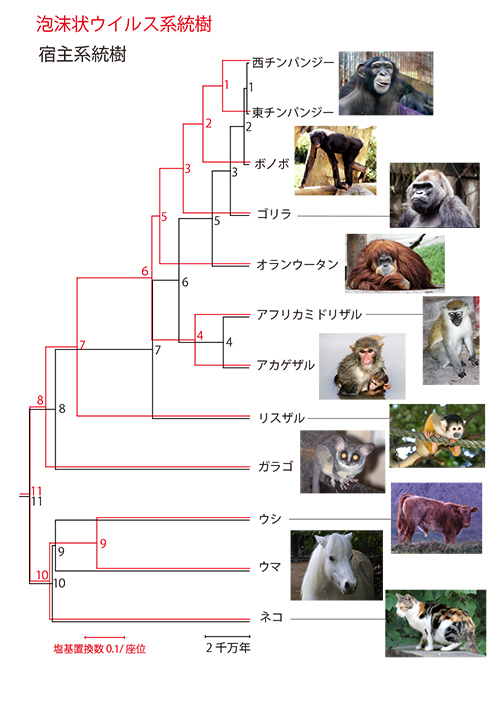
ウイルスの系統樹と宿主の系統樹で全体の幅が一致するように描くと、ウイルス系統樹では現在に近い枝は長い(チンパンジーの二つの亜種であるニシチンパンジーとヒガシチンパンジーや近縁のボノボの枝は長い)が、根元に近い古い枝は宿主にくらべて短くなっていることが分かる。このことをもっと定量的に見るために、図27-5aに分岐年代と泡沫状ウイルスの塩基置換速度推定値の間の相関関係を示す。分岐年代と置換速度の両方を対数で表わすと、きれいな逆相関関係が得られる。
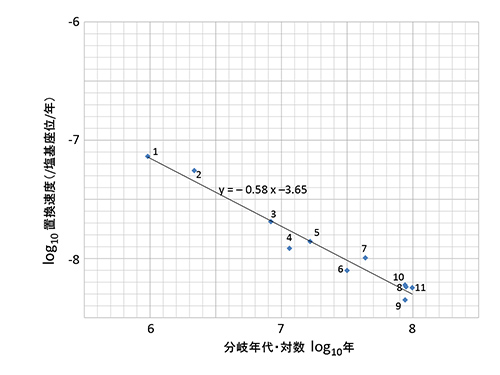
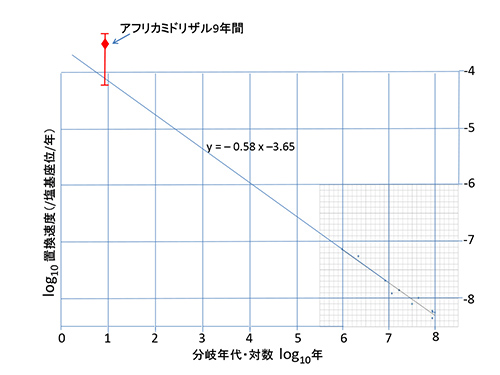
図27-4の節11に対応する霊長目とそのほかのグループが分かれたおよそ9000万年前の分岐から推定される塩基置換速度は5.7x10-9/塩基座位/年となるが、節1に対応するチンパンジーの二亜種間の96万年前の分岐から推定される置換速度は7.3x10-8/塩基座位/年となり、異なる時間スケールで観測されるウイルスの進化速度が1桁以上違うのである。
また、アフリカミドリザルを宿主とするウイルスでの9年間の測定結果によると、突然変異率は3.7x10-4/塩基座位/年とさらに高くなる(4)。
図27-5bは、図27-5aにアフリカミドリザルウイルスのデータを付け加えたものである。ここで付け加えられたプロットは9年間で観測された6回の塩基置換に基づくものであるので、大きな誤差を含む。この6回の置換はたまたま実現したものであり、置換速度が変わらなくても確率的な変動があるので、4回になったり、8回になったりすることもある。
図ではそのような誤差の幅も示したが、誤差を考慮すると9年の時間スケールで得られたアフリカミドリザルのプロットも、96万年~9000万年の時間スケールのプロットから得られた回帰直線の延長上に驚くほどよく乗っていると見なせるであろう。
xを年で測った時間、yを塩基座位当たり年当たりで測った置換速度とすると、9年間の観測で得られた置換速度とそれよりも7桁も大きな9000万年の時間スケールで観測された置換速度が、共通の直線log(y) = -0.58log(x) – 3.65で表現できるのだ。
アフリカミドリザルウイルスの突然変異率の値は、第7話で紹介したコロナウイルスの置換速度に匹敵する。つまり、4年間の時間スケールで測定したMERS-CoVの置換速度7.8x10-4/塩基座位/年、50年間で測定した風邪コロナウイルスHCoV-OC43の2.4x10-4/塩基座位/年とよく一致するのである。
従って第7話で議論したように、コロナウイルス科全体の最後の共通祖先LCAも、泡沫状ウイルスと同じように、1億年スケールの古い時代にまでさかのぼる可能性は大いにあり得ると思われる。
ただし、コロナウイルスの場合は宿主と共進化してきたわけではないので、ウイルスのデータからその分岐年代を高い信頼度で推定することが困難であった。
ところが、泡沫状ウイルスの場合は、時には宿主を換えることはあるものの、基本的には宿主の種分化にあわせてウイルスも種分化してきたことが明らかなので、ウイルスの分岐年代についても信頼性の高い推定が得られるのである。
1億年スケールでは泡沫状ウイルスの塩基置換速度は5.7x10-9/塩基座位/年と推定される。宿主であるヒト上科霊長類の核ゲノムの塩基置換速度は4.3~7.9x10-10/塩基座位/年と推定されている(5)。
またヒトのミトコンドリアのたんぱく質コード領域では、1.2x10-8/塩基座位/年と推定されている(6)。泡沫状ウイルスの置換速度は、短い時間ではほとんどリアルタイムで進化の様子を追跡できるコロナウイルス並みに高いが、1億年に近い時間で見ると宿主の核ゲノムよりは高いが、ミトコンドリアよりは見かけ上低くなるのだ。
時間スケールを長くしていくと見かけ上の置換速度が次第に低くなる理由についてはさまざまなことが考えられるが、このことは泡沫状ウイルスだけでなく、詳しく調べられたたいていのウイルスについて一般的に成り立っているように思われる。第7話で議論したように、多重置換による見かけ上の効果と短時間での弱有害変異の効果とが主要な原因と考えられる。
【引用文献】
1. Katzourakis, A. et al. (2014) Discovery of prosimian and afrotherian foamy viruses and potential cross species transmissions amidst stable and ancient mammalian co-evolution. Retrovirology 11, 61.
2. 長谷川政美(2018)『マダガスカル島の自然史』海鳴社.
3. Aiewsakun, P., Katzourakis, A. (2015) Time dependency of foamy virus evolutionary rate estimates. BMC Evol. Biol. 15, 119.
4. Schweizer, M. et al. (1999) Genetic stability of foamy viruses: long-term study in an African green monkey population. J. Virology 73, 9256–9265.
5. Besenbacher, S. et al. (2019) Direct estimation of mutations in great apes reconciles phylogenetic dating. Nature Ecol. Evol. 3, 286–292.
6. Soares, P. et al. (2009) Correcting for purifying selection: an improved human mitochondrial molecular clock. Amer. J. Human Genet. 84, 740–759.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




