
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第24話
動物進化に寄与したウイルス
文と写真 長谷川政美
◎胎盤の進化
哺乳類は、卵を産むカモノハシ、ハリモグラなどの単孔類、未熟な状態で産んだ子供を袋の中で育てるカンガルー、オポッサムなどの有袋類、それ以外の哺乳類すべてを含む真獣類から成る。このうち有袋類と真獣類のメスは胎盤をもつ。胎盤は子宮の中で育つ胎児と母親がへその緒でつながっているところであり、母体と胎児のあいだの物質交換の場でもある。このような器官の進化とウイルスとは何の関係もなさそうに思われるが、これから見ていくように、最近の研究で胎盤の進化にウイルスが深く関わっていたということが明らかになってきた。
単孔類以外の哺乳類、つまり有袋類と真獣類をあわせて獣類という。獣類の特徴として単孔類のように卵ではなく子供を産むという「胎生」を思い浮かべるひとが多いであろう。
しかし、胎生という出産方法は脊椎動物だけでもおよそ150の系統で独立に進化したといわれている(1)。その多くは、子供が母体の中で育つものの、母親が卵に詰め込んでおいた資源に頼る方式である。
母親が資源を供給し続ける「母体依存型」の胎生は33回、さらに胎盤が進化したのは20回に過ぎない。
このように胎盤を進化させたのは獣類だけではないものの、これが獣類の重要な特徴であることは確かである。一方、単孔類は卵生ではあるものの、母体依存型に近い特徴を進化させているという(1)。
カモノハシのメスが排卵するのは幅4ミリの卵であるが、受精して17日後に産む卵は縦16ミリ、横14ミリになっている。つまり、カモノハシの受精卵は母親から栄養をもらって大きく成長するのである。
単孔類と獣類の共通祖先は、胎生よりも先に母体依存型を進化させたと考えられる。さらにこの共通祖先は、未熟な状態で卵から産まれる子供を母乳で育てられるようになっていた。
◎胎盤で発現する遺伝子はどこから?
ゲノム刷り込み(ゲノムインプリンティング)という現象がある。哺乳類は父親と母親からゲノムをそれぞれ一揃いずつ受け継ぐが、いくつかの遺伝子は片方の親から受け継いだほうだけが発現する。このようにどちらの親から由来した遺伝子かが記録されていることをゲノム刷り込みという。ゲノム刷り込みは、胎盤の形成や子供の成長に関与する遺伝子でよく見られる。
哺乳類の胎盤で発現するゲノム刷り込みを受けている遺伝子15個のうちで、父親由来が10個だという(2)。これはウマとロバの雑種を使って分かったことであるが、胎児と母親を繋ぐ器官である胎盤が、父方の遺伝子の影響を強く受けているのは、何を意味するのであろうか。
このような現象は、父親由来の遺伝子と母親由来の遺伝子のあいだの利害の対立から生じたと考えられる。メスが胎児に振り向けることのできる資源には限りがある。
メスは別の機会に複数のオスを受け入れる可能性があるので、食べ物が乏しいなど条件の悪い年に身籠った胎児に回す栄養は、抑制的になる傾向がある。将来に余力を残しておく方が、生涯のあいだに残せる子供の数が多くなるのだ。
一方、胎児は自分自身が生き残るためになるべく多くの栄養を獲得しようとする。これは「母子の対立」であるが、見方を変えると「父母の対立」でもある。
母子の対立は、父親由来の遺伝子を介して対立しているのである。父親由来の遺伝子にとっては、母親に多少過剰な負担をかけてもなるべく多くの栄養を獲得しようとすることが、自身のコピーを残す可能性を高め、適応的になる。このため、胎児の成長因子は父親由来のものが発現し、成長を抑制する因子は母親由来のものが発現するような選択圧がかかっていると考えられる(3)。
哺乳類の胎盤で発現するゲノム刷り込み遺伝子の多くが父親由来であるのには、このことが関わっているらしい。
◎レトロウイルス由来の遺伝子Peg10
実はゲノム刷り込みを受ける遺伝子の中で、胎盤で発現するPeg10という遺伝子はもともとレトロウイルスの遺伝子に由来しているという(4)。
Peg10という遺伝子の名前は、“paternally expressed gene 10”からきている。つまり父親由来の遺伝子が発現するということである。
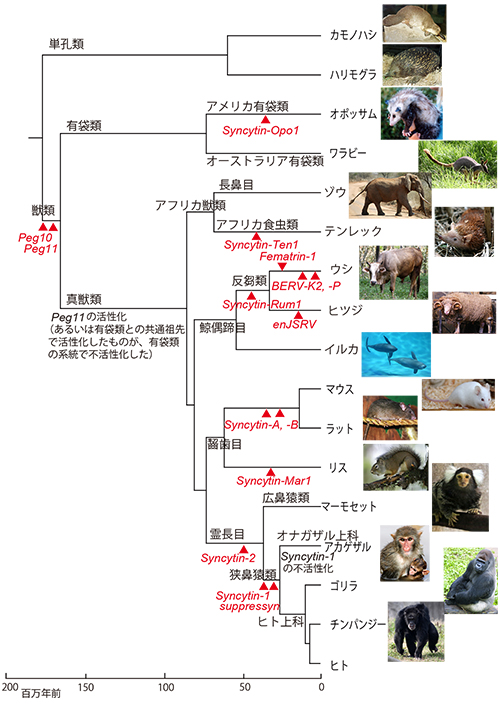
この遺伝子の配列は真獣類と有袋類のゲノム上で保存されており、その周辺の遺伝子も保存されていることから、単孔類と分かれたあとの真獣類と有袋類の共通祖先のゲノムに挿入されたものである(図24-2)。クリックすると拡大表示されます。
遺伝子操作でこの遺伝子を無効化させたマウス(ノックアウトマウスという)では、胎盤形成ができず初期胚は死んでしまう。
これとは別のPeg11/Rtl1 (Rtl1という名前はRetroposon-likeからきている)というやはり胎盤で発現する遺伝子もレトロウイルス由来だが、真獣類でしか保存されていない。
この遺伝子の挿入もPeg10と同様に真獣類と有袋類の共通祖先で起ったが、真獣類でだけゲノム刷り込みを受ける遺伝子として進化し、有袋類では遺伝子としては使われず残骸が残っているだけである(5)。
Peg11/Rtl1をノックアウトしたマウスでは、母親と胎児が栄養やガスのやり取りする胎盤の胎児毛細血管に目詰まりが生じて胎児が死んでしまう(生まれてから死ぬこともあるが)(6)。
◎レトロウイルス由来の「シンシチン」
このように胎盤形成に関わる遺伝子にレトロウイルス由来のものが使われている例は多いが、ここではもう一つ重要なシンシチンを紹介しよう。
胎盤の機能として重要なのが、母親と胎児のあいだの栄養やガス交換の場としての役割だけではなく、子宮の中の胎児を母親の免疫機構による攻撃から守る役割も果たしていることである(7)。
われわれの体内には、病原体から自分を守るために、自分以外の異物、つまり「非自己」を攻撃して排除する免疫機構がある。胎児は母親の遺伝子だけでなく、父親の遺伝子からも作られているので、母親にとっては異物である。従って、子宮の中の胎児を母親の免疫機構による攻撃から守る仕組みが必要である。
胎盤には、絨毛(じゅうもう)を取り囲む「合胞体性栄養膜」という膜がある。この膜は胎児に必要な酸素や栄養は通過させるが、非自己を攻撃するリンパ球などは通さず、胎児を母親の免疫機構による攻撃から守る。
合胞体性栄養膜は細胞が融合してたくさんの核をもった巨大な細胞であり、細胞間の隙間がないために母親の免疫細胞が胎児側に侵入することを防いでいるのである。
実はこの合胞体性栄養膜の形成に重要な役割を果たすシンシチンというたんぱく質が、ウイルスに由来する遺伝子から作られていることが分かったのである。
シンシチンはレトロウイルスがもつenvというエンベロープたんぱく質に由来する。ウイルスの外側を包む脂質二重膜であるエンベロープ(これをもたないウイルスもあるが)は、ウイルスが感染した細胞から飛び出すときに宿主の細胞膜などを剥ぎ取って作るものである。
ウイルス自身の遺伝子の産物であるenvたんぱく質は、このエンベロープに突き刺さるように配置されている。コロナウイルスのスパイクたんぱく質(図2-1)もこれに相当するものである。
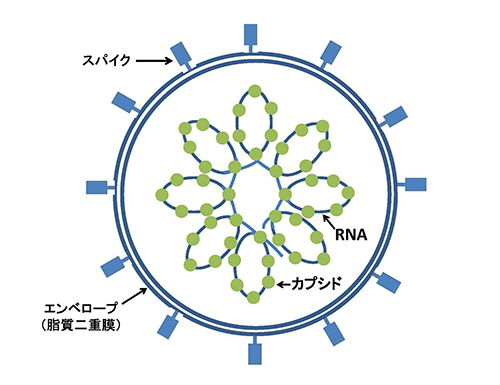
ウイルスが感染する際には、envたんぱく質が感染する細胞の表面にあるレセプターと結合する。その後ウイルスの膜と宿主の細胞膜が融合して、ウイルスは細胞内に侵入する。ウイルスのエンベロープが宿主の細胞膜と同じ脂質二重膜なので、スムーズに侵入できるのであるが、もう一つenvたんぱく質が二つの膜を融合させる橋渡しをしていることも重要である。
このような働きをするレトロウイルスのenvたんぱく質遺伝子がわれわれの祖先のゲノムに取り込まれ、胎盤で発現し、シンシチンを作るようになったのである。シンシチンは細胞融合を起こして、合胞体性栄養膜を作り、胎児を母親の免疫機構による攻撃から守っている。
このようなことが可能になったのは、その起源となったレトロウイルスのenvたんぱく質がもともと細胞膜融合能をもっていたからなのだ(7)。
◎シンシチンはどのように生まれたか
ヒトのシンシチンには2種類ある。シンシチン1とシンシチン2である。前者はヒト内在性レトロウイルスW(HERV-W)、後者はヒト内在性レトロウイルスFRD(HERV-FRD)それぞれのenvに由来するたんぱく質である。これらの内在性レトロウイルスはヒトゲノム中に98,000個も存在し、ゲノム全体のおよそ8%を占める。
シンシチン2は広鼻猿類とヒトを含む狭鼻猿類で保存されており、いずれも細胞膜融合能を保持している。従って、この遺伝子は狭鼻猿と広鼻猿の共通祖先で獲得され、今日まで胎盤形成に重要な役割を果たしてきたと考えられる(図24-2)。
一方、シンシチン1は広鼻猿と分かれたあとの狭鼻猿類の共通祖先で獲得されたと考えられる。ところが、狭鼻猿の中でもヒト上科(ヒトと類人猿)を除いたオナガザル上科ではこの遺伝子に停止コドンやフレームシフトなどの変異が起っていて、不活性になっている。その代わりにオナガザル上科では、内在性レトロウイルスERV-V2のenv遺伝子に由来するmac-Syncytin-3が合胞体性栄養膜の形成に関与している (9)。
シンシチンがヒトの祖先で重要な働きをするようになったのは、真獣類の進化の歴史の中でも比較的最近のことで、狭鼻猿類の段階になってからである。このことは、Peg10が真獣類と有袋類の共通祖先の段階から一貫した役割を果たし続けてきたことと対照的である。
ところが、シンシチンが生まれたのは霊長類の系統だけではなく、マウスとラットの共通祖先、オポッサムの祖先などそのほかにも獣類のさまざまな系統のゲノムに、レトロウイルスのenv遺伝子が独立に挿入され、胎盤で働くようになったのである(図24-2)。
哺乳類の胎盤は非常に多様であり、近縁な種が必ずしも似た胎盤をもっているとは限らない。env遺伝子がゲノムのさまざまな場所に独立に挿入されたことが、この多様性を生み出した原因になった可能性がある。
シンシチン遺伝子は、真獣類の進化の過程でさまざまな内在性レトロウイルスから繰り返しゲノムに取り込まれて胎盤で使われるようになった。
また不活性になって、使われなくなるということもしばしば起こった。このことは、さまざまなウイルスの内在化が継続的に起ることによって、ゲノム中に似た遺伝子配列が複数個共存することを意味する。このような状況では、機能している遺伝子が偶然置き換わってしまうといった進化も起こると考えられる。このような考えは、「バトンパス仮説」と呼ばれている(8)。
このようにレトロウイルスの遺伝子がわれわれの祖先のゲノムに挿入され利用されることがなかったら、安全な母の胎内で十分に成長してから生まれるという現在のヒトは存在しなかったはずである。
【引用文献】
1.リアム・ドリュー(2019)『わたしは哺乳類です ― 母乳から知能まで、進化の鍵はなにか』梅田智世訳、インターシフト.
2.Wang, X. et al. (2013) Paternally expressed genes predominate in the placenta. Proc. Natl. Acad. Sci. USA 110, 10705–10710.
3.Haig, D. (2002) Genomic Imprinting and Kinship. Rutgers Univ. Press.
4.Suzuki, S. et al. (2007) Retrotransposon silencing by DNA methylation can drive mammalian genomic imprinting. PLoS Genet. 3(4), e55.
5.Edwards, C.A. et al. (2008) The evolution of the DLK1-DIO3 imprinted domain in mammals. PLoS Biol. 6 (6), e135.
6.Sekita, Y. et al. (2008) Role of retrotransposon-derived imprinted gene, Rtl1, in the feto-maternal interface of mouse placenta. Nature Genet. 40, 243-248.
7.中屋敷均(2016)『ウイルスは生きている』講談社.
8.Imakawa, K., Nakagawa, S., Miyazawa, T. (2015) Baton pass hypothesis: successive incorporation of unconserved endogenous retroviral genes for placentation during mammalian evolution. Genes Cells 20, 771-788.
9.Esnault, C. et al. (2013) Differential evolutionary fate of an ancestral primate endogenous retrovirus envelope gene, the EnvV Syncytin, captured for a function in placentation. PLoS Genet. 9(3), e1003400.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




