
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第7話
コロナウイルス科の進化
文と写真 長谷川政美
前回までは、ヒトに感染するコロナウイルスの起源を探ったが、今回はコロナウイルス科全体の進化を、その時間スケールに注目して見てみよう。◎コロナウイルスの命名者
1960年代にイギリスの風邪研究所のデビット・ティレルは風邪ウイルスの分離を試みていた。彼は聖トマス病院のアンソニー・ウォーターソンに相談したところ、そこで当時採用されて間もないジューン・アルメイダ(1930~2007)という電子顕微鏡技師が、ティレルの持ち込んだ気管培養組織のサンプルを調べることになった。
彼女が電子顕微鏡でそのサンプルを観察したところ、ウイルスのまわりが王冠のような輪で取り囲まれていることに気がついた。彼女は以前に、ニワトリの気管支炎とマウスの肝炎で似たようなウイルスを観察していた。アルメイダらは風邪のウイルスも含めて、これらのウイルスをラテン語の王冠を意味する「コロナウイルス」と命名し、それを伝える短報がNature誌に掲載された(1)。
この短報では「太陽のコロナ」を彷彿させる、と述べられている。アルメイダが観察した風邪のコロナウイルスは、現在「HCoV-OC43」と「HCoV-229E」と呼ばれているものである。
アルメイダは家庭の事情で大学に進学できず、病院の技師として働きながらキャリアを重ねていた。彼女は電子顕微鏡によるウイルスの画像化に現在でも使われているネガティブ染色という画期的な方法を導入して、「太陽のコロナ」のようなウイルスをはじめて目にしたのだ。
◎進化の時間スケール
コロナウイルス(CoV)科は4つの属に分けられる。アルファコロナウイルス属(α-CoV)、ベータコロナウイルス属(β-CoV)、ガンマコロナウイルス属(γ-CoV)、デルタコロナウイルス属(δ-CoV)である。このうち、α-CoVとβ-CoVは主に哺乳類、γ-CoVとδ-CoVは主に鳥類を宿主としている。
これらさまざまな動物を宿主とするコロナウイルスのゲノム配列を使って系統樹を描くと、図7-1aのようになる。
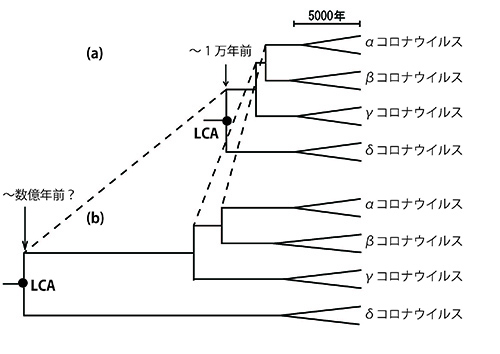
系統樹上の「分岐年代」の推定には、分子進化速度が時間的に一定である「分子時計」を使う。RNAを鋳型としてRNAを合成する酵素である「RNAポリメラーゼ遺伝子」の進化速度(塩基置換速度)がおよそ1.3 x 10-4/塩基座位/年であるという分子時計を基準にして分岐年代を推定するのだ。
そうすると、あらゆるコロナウイルスの「最後の共通祖先」(図の中の「LCA;Last Common Ancestor」)の年代がおよそ1万年前となる。1万年前にまずδ-CoVがほかのグループから分かれ、その後7900年前にγ-CoVが分かれ、7200年前にα-CoVとβ-CoVが分かれたことになる。それぞれの属の中で、さまざまな宿主特異的なCoVが生まれたことになる(2)。
コロナウイルス進化のこの時間スケールは、宿主である哺乳類や鳥類が進化してきた時間にくらべると非常に短いものであるが、コロナウイルスが宿主の種の壁を越えて感染することもあるので、一応は可能なシナリオのように思われる。
コロナウイルスのゲノムの進化速度は、動物にくらべて高いので、確かに数千年から1万年という通常の生物進化の時間スケールからみると短い期間に、多様な進化が起ったように見えるのである。
しかし、α-CoVとβ-CoVはコウモリをはじめとした哺乳類、γ-CoVとδ-CoVは鳥類を主な自然宿主としていることを考えると、多様なコロナウイルスは、宿主である哺乳類や鳥類の進化にあわせて一緒に進化してきた可能性も考えられる。
宿主の種分化にあわせて共生体が一緒に種分化することを「共進化」というが、そのようなシナリオが本当だとすると、数千万年から数億年という進化の時間スケールを考えなければならなくなる。時間スケールが単純な方法による推定値と4桁ほども違っているこのようなシナリオは、コロナウイルスのゲノム配列データと両立し得るのだろうか。あるいは、そのような可能性はコロナウイルスのゲノム配列データからは完全に否定されるのだろうか。実はそのような可能性もありそうなのである(3)。
◎系統樹を古くまでさかのぼると見えること
図7-1aの時間スケールは、コロナウイルス遺伝子の進化速度が1.3 x 10-4/塩基座位/年であるとして推定されたものである。この進化速度は図7-2で示したようにして推定される。感染期のさまざまな期日で採取されたウイルスの遺伝子配列をもとにして描いた系統樹が図7-2だ。
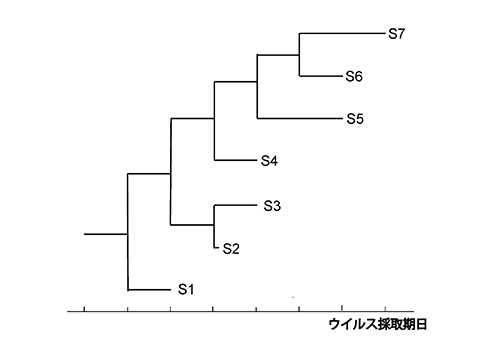
S1のように初期に採取されたウイルスの枝に比べて、S7のように後期に採取されたウイルスの枝は突出している。S1に比べてS7は長い時間進化したので、それだけ多くの変異が蓄積しているからである。このような進化時間の違いと変異量の違いから、進化速度、つまり時間当たりの「塩基置換速度」を推定できるのである。
図7-1aは、このように最近になって採取されたウイルスの遺伝子配列から推定された進化速度が、コロナウイルス科全体で成立していると仮定したものであり、その結果最後の共通祖先LCAがいたのがおよそ1万年前とされたのである。しかし、分子時計の仮定が正しいとしても、この結論が正しいとは限らないことを見ていこう。
最大の問題は、遺伝子の塩基座位によって進化速度は大きく異なることである。たんぱく質をコードする遺伝子では、コドンの3番目(表4-2:3rd)の座位が変わってもアミノ酸は変わらないことが多いので中立的な変異として受け入れられやすい。
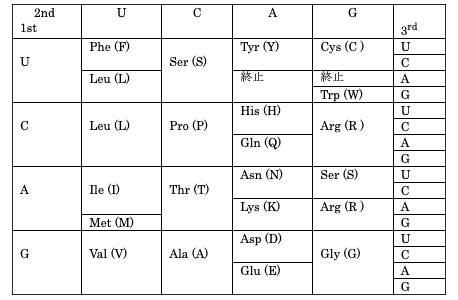
ところが、コドンの1番目(表4-2:1st)あるいは2番目(表4-2:2nd)の座位が変わるとたいていアミノ酸が変わってしまう。そのような変異がウイルスの適応度(感染率や増殖効率など)を高めることも稀にはあるが、たいていの場合は、たんぱく質の変化によってそれまでの機能を果たせなくなってしまう。
この場合、負の自然選択によってそのような変異ウイルスは取り除かれる。つまり塩基の置換速度は座位ごとに非常に違っていて、負の自然選択があまり働かないコドンの3番目など限られた座位に集中的に変異が蓄積しているのである。
このように集中的に変異が蓄積している座位を、同種のウイルス間で比較する場合にはあまり問題にならないが、違う属間など系統的に離れたものを比較する場合には、一つの座位に何回も置換を繰り返した結果しか見えないことになる。一つの座位に繰り返し置換が起ることを「多重置換」という(図7-3)。
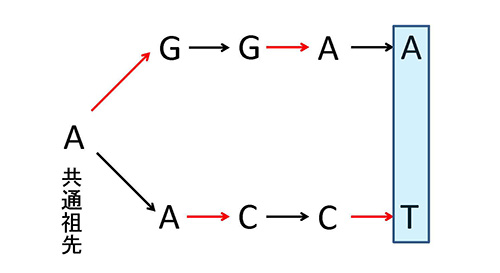
図7-1aでは枝の長さが塩基置換数に比例するように描かれているが、多重置換が多くなるとそれを正しく評価するのが次第に難しくなってくる。このような状況を、「多重置換が飽和している」という。
なるべく現実に近い塩基置換モデルを使って多重置換の効果を評価することが行なわれるが、実際に起こっているウイルス進化の過程は複雑であり、どうしても多重置換の効果が過小にしか評価されない傾向が生じてしまうのである。
このような過小評価の程度は、古い分岐ほど深刻になるから、図7-1bで模式的に示したように、古い分岐ほど比例関係で推定される以上に実際には古くなっている可能性があるのだ。
◎コロナウイルスのLCAは数億年前に存在?
もう一つの問題は、コロナウイルス科の系統樹を古い時代にさかのぼったところにまで、一つの感染期に採取されたウイルスから推定された置換速度を当てはめることである。
第4話で「同義置換」と「非同義置換」の話をした。筆者は以前、霊長類のミトコンドリアDNAの解析をしていた時に奇妙なことに気がついた(4)。非同義置換距離と同義置換距離の比(dN / dS)がヒトとチンパンジーの間など異種間で測った場合に比べて、同種間で測ると5~10 倍くらいになるのである。ここでも多重置換の飽和が影響していることも確かだが、それだけでは説明がつかない。
アミノ酸を変化させる非同義置換はたいていの場合に有害になるが、この違いは、短い時間スケールでみると集団内では少しだけ有害な(弱有害という)変異が蓄積しているためだと解釈できる。弱有害な変異は長い進化の時間スケールでは負の自然選択によって次第に取り除かれるので、種間の比較では dN / dS 比が小さくなるが、種内の比較では最終的に落ち着いた状態ではなく、過渡期をみているために大きくなると考えられるのだ。
同じようなことは、コロナウイルスの進化でも起こっていると思われる。一つの感染期に採取されたウイルスの中には弱有害(あくまでもウイルスが増殖する上で少し不利だという意味)の変異も含まれているはずであり、系統樹をもっとさかのぼるとそのような変異の多くは負の自然選択で取り除かれていると考えられる。
SARSの流行中に感染者16人から採取されたSARS-CoVゲノムの系統樹を描き、置換速度を推定した研究がある(5)。それによると同義置換速度1.67~4.67x10-3/座位/年、非同義置換速度1.16~3.30x10-3/座位/年となる。非同義置換速度は同義置換速度よりも少し低くはなっているが、通常の分子進化ではもっと抑えられていると考えられる。致命的な欠陥でない限りそのような変異でも短期間は存続できているようである。
SARS-CoV-2に関しては、もっと大規模な研究がなされている。COVID-19の流行中に数か月間の感染者1万人余りから採取されたSARS-CoV-2ゲノムの系統樹から同義置換数と非同義置換数を比較したものである(6)。この研究では、1965回の同義置換が観測されているのに対して、アミノ酸を変える非同義置換は2969回であった。このような置換数の比較だけからは、非同義置換が同義置換にくらべてどの程度抑えられているかは分からない。完全にランダムに変異が起こった場合にくらべて、非同義置換数/同義置換数の比がどうなっているかを見なければならない。少し専門的になるので詳細は省略するが、ランダムに変異が起こるとすれば、自然選択の影響を考えないと非同義置換は同義置換にくらべて3.34倍起こりやすいのである。
コドンの3番目の置換以外はたいてい非同義置換であるし、コドンの3番目の置換でも非同義置換のこともあるので、非同義置換のほうが同義置換よりも3.34倍起こりやすいということになる。実際にはCOVID-19の流行中に数か月間で観測された非同義置換数と同義置換数の比は、2969/1965 = 1.51であるから、非同義置換はランダムな場合にくらべて、1.51/3.34 = 0.46の割合で抑えられているように見える。自然選択の影響でこれだけ抑えられているのである。
通常の分子進化ではどうであろうか。583個の遺伝子についてヒトとネズミの間で比較すると、非同義置換速度と同義置換速度の比は、0.14になる(7)。これにくらべると、数か月間のSARS-CoV-2のデータから得られた0.46という値は、3倍以上も大きい。
非同義置換速度はそれぞれの遺伝子に働く機能的な制約の強さによって大きく変わるので、一概には言えないが、このことはコロナウイルスの短い流行期に見られる変異には、弱有害なアミノ酸置換を含むものが多く含まれるが、長い年月にわたる進化でみると弱有害な変異は取り除かれる可能性を示唆するものである。この推測が正しければ、短期間で測られた進化速度にくらべて、長期間で測られた進化速度は低くなることが予想される。
実際のコロナウイルスのデータで、このことを支持する結果が得られている(表7-1)(8)。
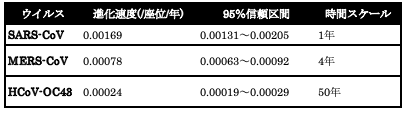
1年程度の時間で測られたSARS-CoVの進化速度、4年程度のMERS-CoV、50年程度のHCoV-OC43がそれぞれ0.00169、0.00078、0.00024/塩基座位/年となり、長期間で測定されたものほど速度が低くなっている。
このように、短い時間スケールで推定された置換速度は、もっと長い時間の置換速度よりも高くなっているようである。このように考えると、コロナウイルス科のLCAはさらに古くなり、数億年前という可能性もあり得るかもしれない。
【引用文献】
1.Almeida, J.D. et al. (1968) Coronaviruses. Nature 220, 650.
2.Woo, P.C. et al. (2012) Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 86, 3995-4008.
3.Wertheim, J.O. et al. (2013) A case for the ancient origin of coronaviruses. J. Virol. 87,7039-7045.
4.Hasegawa, M., Cao, Y., Yang, Z. (1998) Preponderance of slightly deleterious polymorphism in mitochondrial DNA: Nonsynonymous/synonymous rate ratio is much higher within species than between species. Mol. Biol. Evol. 15, 1499-1505.
5.Zhao, Z. et al. (2004) Moderate mutation rate in the SARS coronavirus genome and its implications. BMC Evol. Biol. 4, 21.
6.Koyama, T., Platt, D., Parida, L. (2020) Variant analysis of SARS-CoV-2 genomes. Bull. World Health Organ. 98, 495-504.
7.宮田隆(2014)『分子からみた生物進化』講談社.
8.Boni, M.J. et al. (2020) Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nature Microbiol. https://doi.org/10.1038/s41564-020-0771-4.
9.Hou, W. (2020) Characterization of codon usage pattern in SARSCoV-2. Virology Journal. DOI: 10.21203/rs.3.rs-21553/v2.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。







