
ヒトとチンパンジーの共通祖先は600万年前に生きていた。
この地球上に、ヒトとゾウの共通祖先は9,000万年前、
ヒトとチョウの共通祖先は5億8,000万年前、
ヒトとキノコの共通祖先は12億年前に生きていた。
15億年前には、ヒトとシャクナゲの共通祖先が生きていたという…。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。復旦大学生命科学学院教授(中国上海)。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)『遺伝子が語る君たちの祖先―分子人類学の誕生』(あすなろ書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。

僕たちの祖先をめぐる15億年の旅
第11話
卵を産んでいた僕たちの祖先
文と写真 長谷川政美
◎有袋類との出会い
およそ9,000万年前にさかのぼり、僕たちヒトを含む真獣類の共通祖先に出会うところまで前回お話ししました(図1-2の●13)。その祖先の姿は推測ですが、体長3〜5cmほどのトガリネズミのようだったと考えられています。 今回は、そこからさらにさかのぼります。
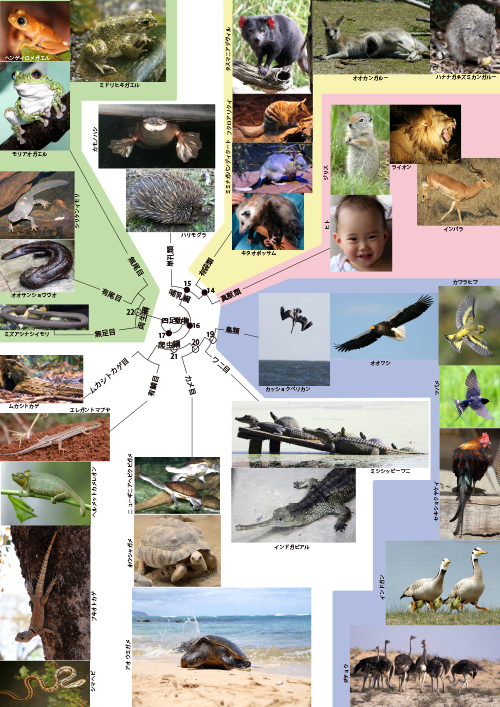
祖先をさかのぼってゆくと、四足動物の系統樹マンダラ(図11-1)でカンガルーなど有袋類との共通祖先●14に出会います。有袋類はメスがおなかに袋をもっていて、そのなかで子供を育てる哺乳類です(図11-2)。

彼らは南アメリカとオーストラリア、ニューギニアに分布しています。キタオポッサムのように北アメリカにもいますが、これはもともと南アメリカで進化したものが、およそ300万年前に陸続きになったときに北アメリカに進出したことがわかっています。
南アメリカの異節類は、ローラシア起源の真獣類の北方獣類、アフリカ起源のアフリカ獣類とくらべると多様性の面で見劣りしますが、その理由は南アメリカが他よりも小さな大陸であったという以外に、ここには有袋類がいて、真獣類が彼らの生態的地位に侵入できなかったということがあります。

南アメリカでは、チラコスミルス(図11-3)のような肉食有袋類が繁栄していましたし、その他にもフォルスラコスという巨大な肉食鳥類がいました。真獣類としては南蹄類という多様な草食獣が栄えたのですが、昆虫食の異節類が現れた以外には肉食真獣類は進化しなかったようです。
ところが、300万年前に陸続きになった北アメリカから真獣類のネコ科動物が進出してくると、有袋類のチラコスミルスは絶滅してしまいました。
◎カンガルーとの共通祖先
オーストラリアでは有袋類が非常に繁栄していますが、カンガルーはその代表で、たくさんの種に分かれています。「カンガルー」という名前はオーストラリア原住民アポリジニの言葉で「跳ぶもの」を意味する「gangurru」からきました。
後足が発達していて、上の写真(図11-4)のように太い尾でバランスをとりながら跳躍することによって、4本足で走るよりも少ないエネルギーで速く走ることができます。カンガルーのオス同士の争いは足蹴りも使う激しいものです(図11-5)。

オーストラリアには、ヒトが持ち込むまでは真獣類はほとんど進出しませんでした。実はオーストラリアでも真獣類の古い化石が見つかっているのですが、ほかの大陸のような真獣類の繁栄は起らなかったのです。
オーストラリアのように乾燥した大陸では、真獣類よりも有袋類が適応しやすかったという説がありますが、真相はよく分かりません。代わりに多様な有袋類が進化しました。

上の写真(図11-6)の手前がフクロオオカミという肉食有袋類で、左奥の真獣類のオオカミとよく似ています。フクロオオカミはオーストラリアやニューギニアに広く分布していましたが、3万年前頃にヒトが進出してくると急速に減少しました。

オーストラリア原住民アボリジニは、ディンゴ(図11-7)というイヌを連れてきたので、ディンゴとの競争もフクロオオカミの減少に拍車をかけました。その結果、ヒトの進出が遅く、ディンゴのいなかったタスマニア島にだけでフクロオオカミは生き残ったために、タスマニアオオカミとも呼ばれます。そのタスマニアオオカミも1936年に動物園で飼育されていた最後の個体が死んで、地球上から姿を消したのでした。
冒頭の四足動物の系統樹マンダラ(図11-1)にある、有袋類のタスマニアデビルも昔はオーストラリア大陸に生息していましたが、現在はかつてのタスマニアオオカミと同様タスマニア島だけに生き残り、絶滅が危惧されます。
オーストラリアでは、このような肉食性以外にもカンガルー、フクロアリクイ、フクロモモンガ、フクロモグラなどさまざまな有袋類が進化しました。このように多様なオーストラリアとニューギニアの有袋類も、6,000万年前頃に南アメリカから陸続きだった南極大陸経由でオーストラリアに移住した祖先から進化したのです。その頃の南極は緑豊かな大陸でした。それが現在のように氷の大陸になったのにも、大陸移動が関係しています。
◎南極大陸と進化の関係
図11-8が示すように、南極大陸はかなりあとの時代まで南アメリカとオーストラリアにつながっていました。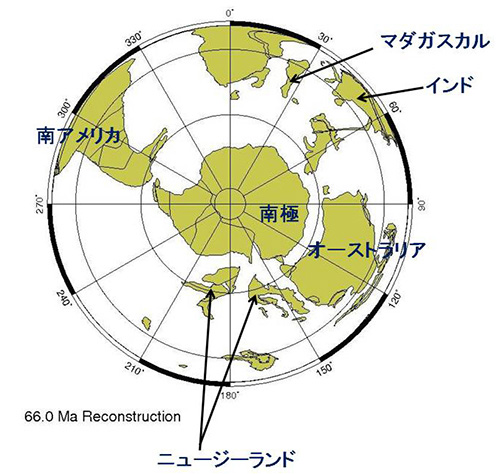
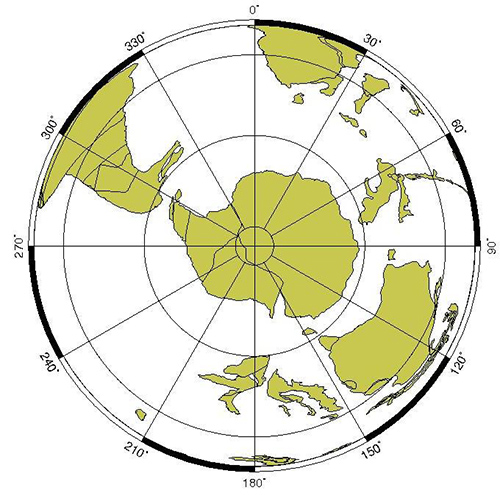
南極が最終的にほかの大陸と分かれて完全に孤立したのが3,500万年前頃で(図11-8b)、それに伴って南極大陸の周りを回る環南極海流が形成されました。そのため、それまで赤道地域から南極大陸沿岸に流れ込んでいた暖流が遮断され、南極は次第に冷えてきたのです。
オーストラリアの有袋類は真獣類のイヌ、ネコ、モモンガ、アリクイ、モグラなどにそれぞれ対応する多様なものに進化したのですが、結局水生適応したものは生まれませんでした。メスが袋のなかで子供を育てるので、そのまま水に入ったら子供が溺れてしまうということがあるからかもしれません。またオーストラリアではカモノハシが水生適応していたので、有袋類が入り込む余地がなかったのかもしれません。
ところが南アメリカでは現地名でヤポックと呼ばれているミズオポッサムが水生適応しています(図11-9)。 
彼らの足には水かきが発達し、水中ではおなかの袋に水が入り込まないよう入り口が閉じるようになっています。僕たちヒトが肛門や尿道口を閉じておくために使う括約筋(かつやくきん)という筋肉がミズオポッサムではおなかの袋を密閉するために使われているのです。こうしてミズオポッサムのお母さんは子供を袋に入れたまま、水のなかを泳ぐことができます。こんな筋肉をうまく使って子供を水のなかにも連れ出せるようにしたとは驚きですね。
◎有袋類は真獣類よりも劣っているか?
ヒトがオーストラリアにディンゴというイヌを連れて来たためにフクロオオカミ(図11-6)が絶滅し、陸続きになった北アメリカから真獣類のネコ科動物がやってきたためにチラコスミルス(図11-3)が絶滅したという話を聞くと、有袋類は真獣類よりも劣っているのではないかと思われるかもしれません。しかし、そんなことはありません。
最初オーストラリアには真獣類と有袋類の両方が進出してきたのに、真獣類のほうは絶滅してしまったという話をしました。また、南アメリカではチラコスミルスのような肉食有袋類がいたために、肉食の真獣類は進化できなかったのです。このように、有袋類が真獣類を圧倒したという歴史もあるのです。
それならばなぜ、フクロオオカミはイヌとの競争に敗れ、チラコスミルスは北からやってきたネコ科動物に敗れたのでしょうか? 一番大きな理由は、オーストラリアや南アメリカが比較的小さな大陸だったからだと思われます。
イヌやネコ科動物は、ローラシア大陸で進化しました。2,000万年前にはアフリカも陸続きになったので、広大な舞台で進化が起ったのです。もちろん、ローラシア大陸といっても、ユーラシアと北アメリカが離れた時期もあります。しかし、重要なことは時々動物相が自由に交流できる時期があったということです。
進化が起った舞台が広いということは、環境も多様だったということです。多様な環境でさまざまな動物と出会う機会が多く、厳しい競争にさらされて進化してきたのが、イヌやネコ科動物だったのです。彼らがオーストラリアや南アメリカに進出して、それまで閉ざされた環境であまり厳しい競争にさらされてこなかったフクロオオカミやチラコスミルスに打ち勝てたのは、当然でしょう。決して真獣類のほうが有袋類よりもすぐれていたからではないのです。
似たような状況は2,000万年前にアフリカがユーラシアと陸続きになった時にも見られました。アフリカから北に進出して成功したゾウや海に進出した海牛目のような例もありますが、ほとんどはアフリカから離れたところにまで分布を広げることはできませんでした。
一方、北の大陸からアフリカにやってきたものの多くが、もともとアフリカで進化した動物に打ち勝ったのです。例えばイワダヌキ目(ハイラックス)は、2,000万年前以前はゾウとならんでアフリカでは主要な草食獣として多様な種類に分かれ、大きさはウサギくらいのものから、スマトラサイくらいのものまでいました。ユーラシアと陸続きになったあと、一時は南ヨーロッパから中国のあたりまで分布を広げましたが、現在ではアフリカと中近東にわずかの種類が残っているだけになり、アフリカでの主要な草食獣の地位は北からやってきたウシ科やウマ科、サイ科などにとって代わられたのです。
また、アフリカにはテンレックの仲間もさまざまな種類がいたのですが、北からやってきた真無盲腸目に押されて、水生適応したポタモガーレ以外は絶滅しました。その前にマダガスカルに渡った仲間は、競争相手がいなくて大繁栄したのですが、アフリカはローラシアとくらべるとやはり小さな大陸だったのです。
このように見てくると、競争に勝つのは真獣類か有袋類かというのではなく、それまでどのような厳しい競争に耐えてきたかが決め手になったのです。
◎カモノハシとの共通祖先
ヒトの祖先をさかのぼるこの旅で次に出会うのが、哺乳類なのに卵を産むカモノハシ、ハリモグラなどの単孔類との共通祖先●15です。 

カモノハシ(図11-10)はオーストラリア、ハリモグラ(図11-11)はオーストラリアとニューギニアに分布しています。
カモノハシがヨーロッパ人によって最初に発見されたのは1798年ですが、この標本を調べた学者は最初哺乳類の毛皮にカモのくちばしを縫い付けたものだと考えたほど、変わった動物でした。その後、人々をもっと驚かしたのは、この動物のメスの腹部にある乳腺から乳が分泌され、それを吸って子供が育つという哺乳類の特徴をもっているにもかかわらず、卵のかたちで子供が生まれてくるという発見でした。
冒頭の四足動物の系統樹マンダラの●15から単孔類、有袋類、真獣類というさまざまな哺乳類が進化してきましたが、僕たちヒトの祖先でもある●15は、カモノハシのように卵を産む動物だったと考えられます。チンパンジーとヒトの共通祖先●1(図2-2 ヒトと類人猿の系統樹)とくらべるとずいぶん遠くまでやって来たものですが、それでもやはり僕たちの祖先なのです。
*もっと詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本連載に大幅な加筆をして、新たな図版を掲載したものです。
扉絵:小田 隆
ブックデザイン:坂野 徹


