
ヒトとチンパンジーの共通祖先は600万年前に生きていた。
この地球上に、ヒトとゾウの共通祖先は9,000万年前、
ヒトとチョウの共通祖先は5億8,000万年前、
ヒトとキノコの共通祖先は12億年前に生きていた。
15億年前には、ヒトとシャクナゲの共通祖先が生きていたという…。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。復旦大学生命科学学院教授(中国上海)。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)『遺伝子が語る君たちの祖先―分子人類学の誕生』(あすなろ書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。

僕たちの祖先をめぐる15億年の旅
第7話
クジラの祖先
文と写真 長谷川政美
◎クジラとカバは仲間
ここでヒトの祖先をたどる旅からは少し横道にそれますが、クジラやウシ、カバなどの進化を見てみましょう。これらの動物の祖先もさかのぼっていくと、いずれヒトの祖先に合流します。
皆さんのなかには、クジラとウシ、カバを一緒のグループにするのはおかしいと思う人がいるかもしれません。確かに以前はウシ、カバ、イノシシ、ラクダなどは蹄(ひづめ)の数が偶数(2本か4本)ということで「偶蹄目(ぐうていもく)」、一方クジラやイルカはクジラ目という別の目に分類されていました。おもに海に住み、陸にあがることのないクジラ類は、陸上で暮らす偶蹄類とは、かたちも生活のしかたもずいぶん違っています。
ところが、DNAを使った分子系統学によって、イルカを含めたクジラ類は、これまで偶蹄目に分類されていた動物のなかで特にカバに近い親戚であることが分かってきたのです。ですから、いわゆる偶蹄目は1つのまとまったグループではなく、クジラ類も含めて「鯨偶蹄目(くじらぐうていもく)」に分類されるようになったのです。
一方、ウマ、サイ、バクなど蹄の数が奇数のものが「奇蹄目(きていもく)」ですが、これらに一番近い親戚がネコ、イヌ、アザラシなどの食肉目とセンザンコウです(図1-2)。
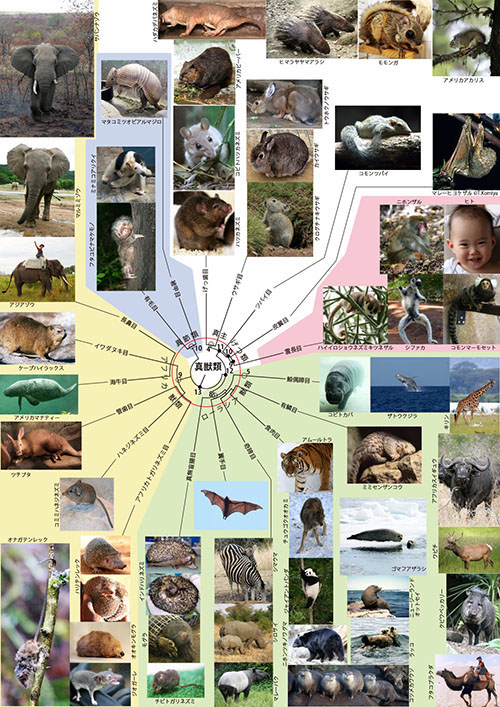
センザンコウは、体毛がうろこ状になっているため「有鱗目」に分類されます。その外見が南アメリカのアルマジロに似ているので、ナマケモノやアリクイとともに「貧歯目(ひんしもく)」に分類されていたのですが、分子系統学によって食肉目の姉妹群として位置づけられるようになりました。
「他人の空似」と言いますが、系統樹上のまったく違うところで、似た動物に出会うことはよくあります。これを「収斂進化」と呼びます。似た環境で似たような生活に適応すると、似たような形態になるということです。陸上から水中の生活に入ったクジラが、サカナと同じような流線形の体型になり、前肢がヒレに変わったのもその一例です。このような例は、この旅のなかで今後もたびたび出会うでしょう。
偶蹄類と奇蹄類は、蹄をもった哺乳類ということで「有蹄類」と呼ばれてお互いに近い親戚だと考えられてきました。蹄は4本の足の先端にある硬い角質の器官で、地面についてからだを支える役割をします。
ゾウも蹄をもつので(図7-1a)、かつては有蹄類に入れられていたのですが、これらのグループはすべて真獣類の系統樹マンダラ(図1-2)の系統樹で離れたところに位置づけられています。

図7-1bは、南アメリカにいてヒトがやってきた後で絶滅したと考えられるトキソドンという哺乳類ですが、これも蹄をもっています。この仲間は「南蹄類」と呼ばれ、その蹄は独自に進化したものと思われます。

どういうことかというと、かつては分類の重要な要素と考えられていた「蹄」も収斂進化で生じたものなのです。蹄をもっているからといって、必ずしも近い親戚だとは言えないのです。
蹄は、速く走って捕食者から逃れるために進化したと考えられます。もちろん捕食者のほうも速く走らなければなりませんが、前肢の爪は獲物をたおすために重要なので、これを蹄に変えることはできなかったのでしょう。また、蹄では足音が響いて、すぐに獲物に気づかれてしまいます。
◎繁栄するコウモリの仲間は?
真獣類の系統樹マンダラ(図1-2)では、奇蹄目+食肉目+有鱗目、鯨偶蹄目、それに翼手目(コウモリ)の3者が同時に分かれたように描かれています。翼手目はかつて、霊長目に近い仲間の主獣類に分類されていましたが、このグループから外されたという話を前にしました。そのコウモリは、実はこんなところに位置づけられたのです。
コウモリの翼は、前腕と指の骨が長くのびてそこに皮膜が張られたもので、膜は後肢までのびています(図7-2)。

よく見ると、膜には細い血管網が張り巡らされていて、翼の激しい運動に必要な代謝を支えています。空を飛べるということでコウモリは非常に繁栄しており、翼手目はげっ歯目に次いで哺乳類のなかで最も種数の多いグループなのです。
系統樹で3つの系統が同時に分かれたように描かれているのは、実際に同時に分かれたという意味ではなく、分かれた順番がはっきりとは分からないということです。
東京工業大学の西原秀典さんと岡田典弘さんが進められ僕も参加した2006年の研究では、「奇蹄目+食肉目+有鱗目」の姉妹群が翼手目であり、鯨偶蹄目はさらにそれらの姉妹群であるという証拠が得られました。しかし、その後の研究でその仮説を補強する結果が得られないので、この問題はまだ決着がついていないということです。
奇蹄目+食肉目+有鱗目と翼手目が一つのグループを構成するということで、われわれはこのグループを「ペガソフェラエ」と名づけました。ウマと空を飛ぶコウモリを含むということでギリシャ神話の空飛ぶウマの「ペガサス」、それに食肉目+有鱗目を「フェラエ」と呼ぶのでこの2つをあわせて命名したのです。
これに対して、「奇蹄目+食肉目+有鱗目」の姉妹群が鯨偶蹄目であり、翼手目はさらにそのグループの姉妹群だという説もあって、「奇蹄目+食肉目+有鱗目」、翼手目、鯨偶蹄目の3者の間の系統関係はまだ決着していないので、同時に分かれたように描かれています。この3者間の枝別れは、順番をはっきりと決めるのが難しいくらい地質学的には短期間に続いて起ったことは間違いありません。
「奇蹄目+食肉目+有鱗目」、翼手目、鯨偶蹄目の3者が一つのグループを構成しているということはほぼ確かで、その姉妹群がハリネズミ、モグラ、トガリネズミなどの「真無盲腸目」です。
無盲腸目は、以前食虫目とも呼ばれていましたが、食虫目にはマダガスカルのテンレックやアフリカのキンモグラなども含まれていました。ところが分子系統学はテンレックやキンモグラを別のところに位置づけたために、これらを除いたものが真無盲腸目として分類されるようになったのです。ここで除かれたテンレックとキンモグラとは、今回の旅のあとでまた出会うことになります。
.jpg)
こうしてカバ、クジラ、ウシ、センザンコウ、トラ、ウマ、コウモリ、ハリネズミなどの祖先をたどっていくと、共通祖先○8に出会います。この祖先から進化した鯨偶蹄目、有鱗目、食肉目、奇蹄目、翼手目、真無盲腸目をあわせて「ローラシア獣類」と呼びます。ローラシアとは中生代に北半球にあった大陸の名前で、○8はそのローラシア大陸で進化したものと考えられています。
◎クジラとカバの共通祖先
ここで鯨偶蹄目について、もう少し詳しく見てみましょう。図7-3が鯨偶蹄目の系統樹マンダラです。

ウシ科、シカ科、プロングホーン科、キリン科、それにマメジカ科が1つのグループを作っていますが、これが反芻類です。セルロースの多い植物食であるため、胃がいくつかの部屋に分かれていて、そのなかの1つでバクテリアにセルロースを消化してもらっています。
反芻類の姉妹群がクジラ類+カバ科です。つまりクジラはカバと共通の祖先から進化したのです。クジラが陸上の真獣類から進化したものであることは以前から認められていましたが、カバがクジラに一番近い真獣類であることをはじめて示したのは、東京工業大学の二階堂雅人さんと岡田典弘さんによる分子系統学的な解析でした。
カバは水中にいることが多いので、カバとクジラが近い親戚であることに納得する人は多いかもしれませんが、カバからクジラが進化したわけではありません。カバとクジラが共通の祖先(図7-3の○5)から、それぞれ進化したということです。
.jpg)
共通祖先○5が、現在のカバのように多少とも水生適応したものであった可能性は低いと思われます。クジラの祖先と考えられる化石は5,300万年前のパキケタスまでさかのぼることができるのに対して、カバの祖先と考えられる化石は1,000万年前くらいまでしかさかのぼれないのです。ですから、クジラとカバの共通祖先がどのようなものであったかを知る手掛かりがありません。
また、現生のカバは昼間水中で過ごすことが多いのですが(図7-4)、彼らは決して泳ぎがうまいとは言えません。カバももちろん泳ぎますが、水中にいる時間の多くは、浅瀬でじっとして過ごしているのです。

5,300万年前にクジラの直接の祖先がいたということは、クジラとカバの共通祖先はそれよりも古かったということですが、現在のカバの水生適応の程度からは、そんなに古い時代からずっと水生適応していたとは考えにくいのです。
クジラは非常に特殊化してしまったため、特にカバに近い親戚であることを示す形態的な特徴は見つかりません。ただし、クジラの祖先が陸上にいた頃には、偶蹄類と共通した特徴をもっていたことが分かっています。それは後肢の踝(くるぶし)にある距骨(きょこつ)の二重滑車構造です(図7-5)。

この構造は偶蹄類一般に見られます。もちろん現在のクジラには後肢がありませんから、このような構造はありませんが、同じ構造がムカシクジラのパキケタスにも見られることから、クジラが偶蹄類の仲間であることが分かります。二重に滑車の構造があるので、足首を回せない代わりに前後方向に固定することで速く走るのに適応したと言われています。
偶蹄類ではブタ、ラクダ、ウシ、シカ、カバすべてに共通する特徴であり、速く走るために偶蹄類、正確には鯨偶蹄類の祖先が獲得した形質と考えられます。共通祖先○5の後肢にもこのような構造があったはずです。
クジラには歯をもった歯クジラ(図7-6a)と歯をもたずに上顎から生えた「ひげ」を使ってプランクトンや小魚など小さなエサを濾しとって食べるヒゲクジラ(図7-6b)がいます。イルカは歯クジラですが、マッコウクジラ以外の大型のクジラ類はみなヒゲクジラです。


鯨偶蹄目のなかで最も種数の多いのがウシ科です。ウシ科は、ウシ亜科、ヤギ亜科、ブルーバック亜科など7つの亜科から構成されています。ウシ亜科を構成している属の数はとても多いので、亜科と属の中間段階の分類単位として「族(ぞく)」が設けられています。例えば、ウシ亜科は、ウシ族(ウシ、バンテンなど)、ネジレヅノレイヨウ族(グレータークーズーなど)、ニルガイ族(ニルガイ)の3族から成り立っています。図7-3は鯨偶蹄目における族以上の分類単位の間の関係を描いた系統樹マンダラなのです。
*もっと詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本連載に大幅な加筆をして、新たな図版を掲載したものです。
扉絵:小田 隆
ブックデザイン:坂野 徹


