
ヒトとチンパンジーの共通祖先は600万年前に生きていた。
この地球上に、ヒトとゾウの共通祖先は9,000万年前、
ヒトとチョウの共通祖先は5億8,000万年前、
ヒトとキノコの共通祖先は12億年前に生きていた。
15億年前には、ヒトとシャクナゲの共通祖先が生きていたという…。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。復旦大学生命科学学院教授(中国上海)。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)『遺伝子が語る君たちの祖先―分子人類学の誕生』(あすなろ書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。

僕たちの祖先をめぐる15億年の旅
第10話
恐竜の絶滅と真獣類の進化
文と写真 長谷川政美
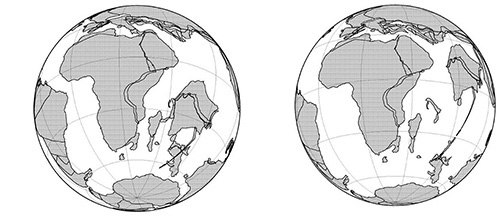
僕たちの仲間の「北方獣類」、ゾウなどの「アフリカ獣類」、ナマケモノなどの「異節類」、これらの3グループのあいだの枝別れは、1億3,000万年前から現在にいたる大陸の分断の歴史(図5-2)と関係があると考えられています。
およそ3億年前から2億年前までのペルム紀と三畳紀には、地球上の大陸はパンゲアと呼ばれる1つの超大陸としてまとまっていました。それが、次のジュラ紀の後期になると、1億5,000万年前までには、北半球のローラシア大陸と南半球のゴンドワナ大陸とに分裂しました。ゴンドワナ大陸は1億3,000万年前頃から分裂を始め、まずマダガスカルとインドの塊がアフリカから分かれました(図5-2a)。
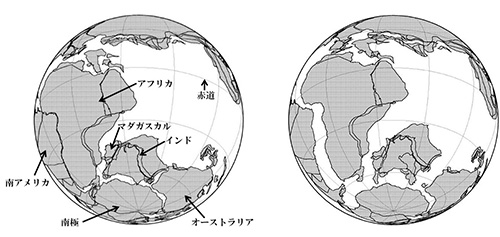
1億500万年前にアフリカと南アメリカとが分裂し(図5-2b)、アフリカはその後2,000万年前にユーラシアと陸続きになるまでは孤立した大陸だったのです。この時期、その大陸で「アフリカ獣類」は進化しました。
一方、アフリカと分かれたあとの南アメリカも、およそ300万年前に北アメリカと陸続きになるまでは、ほぼ孤立した大陸でした。ただし、図5-2から分かるように南アメリカから南極を経由してオーストラリアに至る経路はあとの時代まで保たれていました。この経路は、あとで出てくるように有袋類の進化にとって重要でした。
南アメリカでの「異節類」の進化は、1億500万年前にこの大陸がアフリカから分かれて300万年前に北アメリカと陸続きなるまでのあいだに起ったのです。
僕たちの仲間の「北方獣類」は、南半球のアフリカ獣類や異節類とは別に、北半球のローラシア大陸で進化したと考えられます。
◎3大グループの共通祖先は?
大陸の分断が真獣類の進化と直接結びついているとすると、まずローラシア大陸の北方獣類が、1億5,000万年前までにアフリカ獣類+異節類から分かれて独自の進化の道を歩み始めたということになります。ところが、この年代は、真獣類の進化の歴史を考えると古過ぎます。
また、分子系統学からは、「北方獣類」「アフリカ獣類」「異節類」の3大グループのあいだの枝別れの順番が決められないので、3者間の枝別れは、地質学的には短時間に続けて起ったと思われます。ですから、1億5,000万年前までにまず北方獣類がほかの真獣類から分かれ、4,500万年の長い時間を経て、1億500万年前に南アメリカとアフリカが分裂したのを契機にアフリカ獣類と異節類とが分かれたというシナリオは成り立たないのです。それでは真相はどうだったのでしょうか?
図5-2bをご覧ください。アフリカと南アメリカが分かれた1億500万年前の大陸の配置です。これよりもはるか前にゴンドワナ大陸から分かれたローラシア大陸は、現在のヨーロッパのあたりでアフリカに近いところに位置しています。つまり、ローラシア大陸はゴンドワナ大陸と分かれてはいたものの、長いあいだかなり近いところにとどまっていたのです。従って、これまで新世界ザルやマダガスカルのキツネザル類の起源に関連して出てきたように、浮島による大陸間の移住ということがあり得たはずです。
また、小さな動物の場合は、風に乗って運ばれた可能性もあります。この頃は恐竜全盛時代で、真獣類の祖先は恐竜があまり活動しない夜間だけ活動する小動物だったと思われます。このように小さな動物が強風に乗って海を渡った可能性も考えられるのです。小さな哺乳類のなかには冬眠するものもいるので、仮死状態で風に乗って別の大陸に移住することができたかもしれません。
ローラシア大陸とゴンドワナ大陸のあいだの動物相の交流がこのようなかたちで時々は可能だったと考えれば、「北方獣類」「アフリカ獣類」「異節類」のあいだの枝別れがほとんど同時に起ったように見えることが説明できます。
さらに、この3大グループの枝別れに関連して、もう1つ問題があります。それは最新の分子系統学の研究によると、アフリカ獣類と異節類が分かれたのは、大陸分断の1億500万年前よりも新しいのではないかというのです。およそ9,000万年前という年代が現在一番信用できそうです。
この矛盾も海を越えた移住が、大陸が分かれたあともしばらくは続いたと考えれば説明できます。アフリカと南アメリカの距離がその後次第に大きくなると、なかなか簡単には移住できなくなってきます。それでも何千万年かのあいだには、新世界ザルや新世界ヤマアラシのように移住に成功するものがでてきます。このような移住は、大陸が陸続きになったあとで起る移住とはまったく異なります。
およそ2,000万年前にアフリカがユーラシアと陸続きになったあとでは、北方大陸からウシ科、キリン科、ウマ科、ネコ科などが大挙してアフリカに押し寄せてきました。そのなかのウシ科は現在世界のどの地域よりもアフリカで繁栄しています。鯨偶蹄目の系統樹マンダラ(図7-3)のウシ科動物のなかでアフリカのものが半数近くを占めています。

一方、偶然に頼る浮島などによる移住では、それに成功するグループは限られます。アフリカから南アメリカに移住できた真獣類は、新世界ザルや新世界ヤマアラシの祖先だけだったのです。現在の南アメリカにはジャガーやピューマなどのネコ科動物がいますが、彼らは300万年前に北アメリカが陸続きになったあとで北からやって来たものです。実はそれ以前に、南アメリカには多様な真獣類や有袋類が進化していたのですが、北からの真獣類の移住が始まってから、多くのグループが絶滅に追いやられてしまいました。
大陸の分断が種の分岐に対応するという考えは、理路整然としていて、科学的な検証が可能な仮説としての体裁を整えているために、人気があります。一方、偶然に委ねられた海を越えた移住は直接の証拠を示すことが難しい仮説です。しかし、DNAから推定された系統関係や分岐年代が、大陸分断仮説に合わない場合は、海を越えた移住仮説を考慮せざるを得ないのです。
◎どうして恐竜は絶滅したか
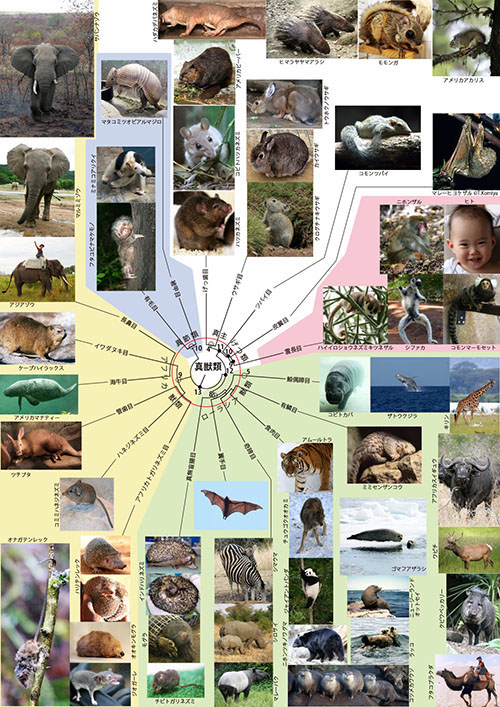
真獣類の系統樹マンダラ(図1-2)の中心部の赤い円は、恐竜が絶滅した6,600万年前に対応します。これは中生代白亜紀と新生代古第三紀の境界に相当するので「K-Pg境界」(白亜紀のドイツ語「Kreide」の頭文字「K」と古第三紀の英語「Paleogene」の略字「Pg」)と呼ばれています。赤い円の内側に分岐が沢山あるということは、真獣類が6,600万年前よりも前にいくつかの系統にすでに分かれていたことを示しています。
恐竜が絶滅した原因は、6,600万年前に巨大隕石が地球に衝突しことが原因であると考えられています。この説は、1980年に地質学者のウォルター・アルバレスとその父で物理学者のルイス・アルバレスが唱えました。彼らは、イタリアにあるK-Pg境界の粘土層に異常に高濃度のイリジウムが含まれていることを見つけたのです。その後、世界各地のK-Pg境界の地層からも高濃度のイリジウムが見つかりました(図10-1)。

この元素は地球の地殻にはほとんど存在しないことから、彼らはこれが隕石の衝突によってもたらされたものであると考えました。そして、恐竜をはじめとしたこの時期の大量絶滅の原因を、イリジウムをもたらした巨大隕石の衝突に求めました(図10-2)。

◎みんなトガリネズミのようだった
恐竜が実際にはどのような原因で絶滅したかは別として、彼らが6,600万年前に絶滅してしまったことは確かです(ただし、恐竜の子孫が鳥類として生き延びているのでこの言い方は厳密には正しくありませんが、そのことについては第12話で触れます)。以前は、恐竜絶滅がきっかけとなって、それまで中生代を通じて恐竜によって占められていた生態的地位(ニッチェ)が空き、それを埋め合わせるべく新生代に入ってから真獣類の急速な進化が始まったと考えられていました。
1つの祖先から多様な系統にいっせいに進化することを、「適応放散」と呼びます。確かに化石記録を見ると、霊長目、げっ歯目、食肉目、鯨偶蹄目、奇蹄目、翼手目、長鼻目などの現在の目(もく)に対応すると思われる動物の化石はすべて6,600万年前よりもあとの時代にならないと見つからないのです。
ですから、恐竜絶滅が引き金となって真獣類の適応放散が起ったように見えますが、唯一の例外が食虫目です。トガリネズミなど食虫類に似た動物の化石は中生代白亜紀の地層からも発見されています。しかし、ハリテンレックやキンモグラのところで述べたように、食虫目は現在では進化的に1つのグループとは見なされなくなってきました。恐竜全盛の時代においては、真獣類はすべてトガリネズミに似た夜行性の食虫類的な動物であったと考えられます。哺乳類にはそれ以外の生態的地位は残されていなかったのです。ところが、大陸分断などによって、遺伝的分化は進んでいたと考えられます。
真獣類の系統樹マンダラ(図1-2)で、恐竜絶滅以前から真獣類の分化が進んでいたように描かれているのは、あくまでも遺伝的分化が進んでいたということであって、形態は必ずしも現在生きている子孫に似たものではなかったはずです。僕たちヒトの祖先である●9、●10、●11や、グリレス類の共通祖先○4は、6,600万年前よりも古い中生代白亜紀に生きていましたが、それらはみな現在生き残っている子孫よりはトガリネズミに似た食虫類的な動物だったと思われます(図10-3)。恐竜が絶滅して初めて現在の目に対応するような形態的な進化が可能になったのです。

*もっと詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本連載に大幅な加筆をして、新たな図版を掲載したものです。
扉絵:小田 隆
ブックデザイン:坂野 徹


