
はるかむかし地球上のあらゆる生物の共通の祖先がいた。
まだ見ぬその姿は細菌のようだったのだろうか?
ダーウィンが夢見た、共通祖先から連なる全生物の進化の系統樹が、
生物のもつDNA情報にもとづいて、いまその姿を現しつつある。
進化生物学者である著者が撮影した貴重な写真コレクションを用いて、
動物界、なかでも昆虫綱、哺乳綱、鳥綱の驚きの進化を100点超の系統樹で表現する。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。最新刊は『ウイルスとは何か』(仮題、中公新書、2023年1月刊行予定)。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

すべての生き物をめぐる
100の系統樹
第16話
ヤガ上科の系統樹マンダラ
文と写真 長谷川政美
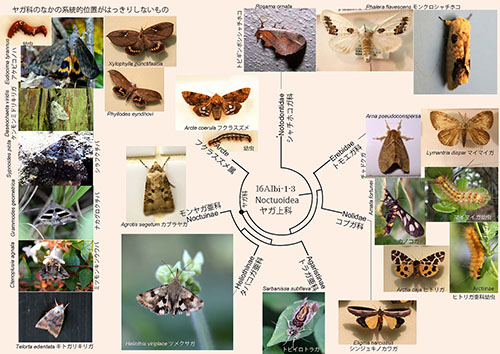
◎鱗翅目のなかで最大の上科
ヤガ上科Noctuoideaは鱗翅目のなかで最大の上科であり、4万以上の種を含む。このなかで最大の科がトモエガ科(およそ2万5000種)、次がヤガ科(およそ1万2000種)である。
ヤガ上科にはいわゆる「害虫」と見なされているものが多いため、農業、林業や環境衛生上の観点から多くの研究がなされている。それに伴い、たくさんの種の分子系統樹解析もなされているが、膨大な数の種のなかで解析されているのはごくわずかであり、まだ系統的な位置づけがなされていないものも多い。
図16AIbi-1-3のなかには、近縁と思われる種について分子系統解析がなされている場合に、近縁性を仮定して系統樹上に位置づけたものもあり、将来この位置づけが訂正されるものもあるだろう。
系統樹マンダラの左に、「ヤガ科のなかの系統的位置がはっきりしないもの」をまとめて示した。文献(6)では日本産のヤガ科だけでも38亜科に分類しているが、私が調べた範囲では、この系統樹に現れる種ではそのうちわずか4つの系統しかはっきり分からないということである。
◎目玉模様のアケビコノハ
系統的位置がはっきりしないもののなかでいちばん上のアケビコノハは派手な目玉模様をもつ(下にも写真で示した)。アケビコノハの後翅の目玉模様は、休んでいるときには普段は枯れ葉に似た前翅に隠れていて見えないが、鳥などに襲われそうになるとこれを露出させ、相手を怯ませて捕食を免れる効果があるとされている(前連載の『進化の目で見る生き物たち』の第25話)。捕食者を驚かせて一瞬でも攻撃を遅らせることが、生死を分ける効果があるのだ。


◎マイマイガと共生細菌
トモエガ科にはチャドクガやマイマイガなど毒針毛をもつものがたくさん含まれる。下の写真は、マイマイガの幼虫であるが、終齢に近いものと思われる。

マイマイガは北アメリカ以外の北半球に広く分布していたが、19世紀後半に北アメリカにも人為的に持ち込まれた。アメリカでは大発生すると森林の樹木の葉を食い荒らすなど深刻な影響を与えている。もともと分布していた地域には寄生バチや病原菌などの天敵がいるために大発生が起きても自然に治まるが、新しく導入されたところでは手に負えなくなるのだ。
それでも、大発生はもともと分布していた西シベリアでもやはり大きな問題である。そこでも時々大発生が起るが、大発生地域の先端が北東方向に1年間で100~200kmくらいの速度で移動するのだという。マイマイガのメスの移動能力は限られているので、そんなに速く大発生が広がるのは何故か、と考えた研究者が面白い仮説を提唱している(8)。
西シベリアのおよそ1400kmにわたる地域のマイマイガのミトコンドリアDNAを調べた結果、地域による遺伝的な違いは見られず、広い地域で遺伝的に均質であることが明らかになった。そこで研究者が考えた仮説は、マイマイガの1齢幼虫が風に乗って空を飛んで移動するというものである。
マイマイガの幼虫は糸を吐いて木からぶら下がることから「ブランコ毛虫」とも呼ばれる。そして本連載の第10話で紹介した空を飛ぶクモのように、風に乗って空を飛んで移動するというのである。確かに大発生の移動方向に風が吹くことが多いという。
日本のマイマイガには、本州、四国、九州に分布するLymantria dispar japonicaと北海道に分布するLymantria dispar praeteraeという2つの亜種がいる。このなかで北海道の亜種に、メスの子供しか育たないメスの系統が見つかっている。このようなメスが産む卵のおよそ50%しか孵化しない。つまり、オスになるはずの卵は発生の途中ですべて死んでしまうのだ。
このメスの子供しか育たない系統を調べたところ、ミトコンドリアは北海道亜種L. d. praeteraeのものではなく、本州亜種L. d. japonicaのものだということが分かった。つまり、2つの亜種の交雑で生まれたものであり、本州亜種のメスからミトコンドリアを受け継いでいるものである。
このようにメスの子供しか育たないような系統は、昆虫ではよく見られる。例えば寄生バチなどでは、メスがボルバキア(Wolbachia)という真正細菌に感染すると単為生殖で繁殖できるようになることがある。このことは、共生細菌によって仕向けられていると考えられている。ボルバキアは母親を通じてしか次世代に受け継がれないので、オスはこの共生細菌の繁殖に貢献しない。宿主が単為生殖するようになれば、生殖のためにオスは必要なくなり、寄生者のボルバキアにとっては都合がよい。オスの存在はボルバキアの増殖に貢献するメスの餌を奪うものだから好ましくないのだ。
しかし、どの動物も単為生殖できるわけではないので、ボルバキアは別の戦略を採ることもある。テントウムシやスジマダラメイガなどでは、ボルバキアは感染したオスを殺してしまうのだ。孵化する前や若齢幼虫の時期にオスだけを殺すという。共生細菌にとっては、オスの子供を早い時期に殺してしまうことが、その分メスの子供の食料を増やし、自分たちの繁殖に貢献することになる(9)。
北海道のマイマイガの亜種間雑種で見られるオスになるはずの卵が発生の途中ですべて死んでしまうという現象は、これに似ている。そこで研究者がボルバキアのような共生細菌がいないか徹底的に調べたが、見つからなかった(10)。マイマイガの亜種間雑種でなぜオスの子供が育たないかは謎である。
その後、北海道の亜種間雑種ではないが、本州亜種でスピロプラズマ(Spiroplasma)という共生細菌が見つかっている(11)。この細菌ではボルバキアと同じように、フキノメイガ(Ostrinia zaguliaevi)というメイガ上科ツトガ科のガに感染すると、オス殺しをすることが知られている。本連載の第1話でお話ししたように、生物進化の初期から異なる系統の生物が共に生きるという共生が重要な役割を果たしてきた。マイマイガの亜種間雑種の謎に共生微生物(ウイルスを含む)が関与しているという証拠は得られていないが、共生はそれが成立している状況に応じてさまざまな展開を見せるという意味で興味深い研究対象である。
◎見事な擬態
下の写真はモンシロドクガかあるいはリンゴケンモンと思われるガの幼虫であるが、この2種は非常によく似ていてこの写真からはどちらなのか判定できない。

図16AIbi-1-3でアケビコノハの下にあるケンモンミドリキリガは地衣類の生えた木の幹にとまっていると、周りに紛れて目立たない。このように目立たないことも身を守る戦略の一つである。その際に手の込んだ方法もある。
今回最初に取り上げたヤガ科のアケビコノハは枯れ葉のように見えるが、シャチホコガ科にも下の写真のトビギンボシシャチホコのように枯れ葉のように見えるものが多い。

ここで、「戦略」とか「工夫」という言葉を使ったが、もちろんガが頭で考えて擬態を設計しているわけではない。ランダムに起るさまざまな変異のなかで、少しでも外敵を欺くことに成功した変異をもった個体が生き残り、さらに変異を蓄積した結果として、見事な擬態が生まれたのである。
◎小コウモリのエコロケーションを混乱させるガ
トモエガ科のガには毒をもったものが多いが、下の写真の南アメリカに分布するヒトリガの仲間Cycnia teneraの成虫も有毒である。

夜行性のガにとって毒をもつだけでは捕食者に対して警告の効果を発揮することは難しい。Cycnia teneraはコウモリに対して超音波を発することで、「自分は有毒だから食べないで」という警告を発するようになった(13)。そのような超音波を発するガを食べたコウモリは、有毒なガを避けるような学習するのだ。ところが、同じような超音波を発する無毒のガも現れているという(14)。これもベイズ型擬態の一種であろう。
◎農業害虫ハスモンヨトウ
下の写真は、農業害虫として悪名高いハスモンヨトウの幼虫(a)と成虫(b)である。多くの昆虫の幼虫は特定の植物しか食べないが、このガの幼虫は40もの科にわたる植物の葉を食べる。

 (b)
(b)
ハスモンヨトウの幼虫はタバコの葉も食べるが、これにはニコチンという有害物質が含まれている。そもそもタバコがニコチンをもつのは、動物に食べられるのを防ぐためだった。従って、これを食べる幼虫は有害物質を無毒化する機構をもっていなければならない。
多くの植物の葉には有害物質が含まれるので、ハスモンヨトウの幼虫のように色々な食草をもつためには、それぞれの植物のもつ多様な毒に対処しなければならず大変なので、たいていの幼虫は限られた種類の植物しか食べない。
最近このハスモンヨトウのゲノムが解読され、カイコの幼虫のようにクワしか食べないようなものに比べると解毒のための酵素の遺伝子が重複して大幅に増えていることが明らかになってきた(15)。また、殺虫剤に対する耐性獲得の機構も次第に解明されつつある。
つづく
【引用文献】
1. Regier, J.C., Mitter, C., Mitter, K., et al. (2017) Further progress on the phylogeny of Noctuoidea (Insecta: Lepidoptera) using an expanded gene sample. Syst. Entomol. 42, 82–93.
2. Keegan, K.L., Rota, J., Zahiri, R., et al. (2021) Toward a stable global Noctuidae (Lepidoptera) taxonomy. Insect Syst. Div. 5(3)
3. Wang, H., Wahlbergc, N., Holloway, J.D., et al. (2015) Molecular phylogeny of Lymantriinae (Lepidoptera, Noctuoidea, Erebidae) inferred from eight gene regions. Cladistics 31, 579–592.
4. Ghanavi, H.R., Twort, V., Hartman, T.J., et al. (2021) The (non) accuracy of mitochondrial genomes for family level phylogenetics: the case of erebid moths (Lepidoptera; Erebidae). bioRxiv
5. Zahiri, R., Holloway, J.D., Kitching, I.J., et al. (2012) Molecular phylogenetics of Erebidae (Lepidoptera, Noctuoidea). Syst. Entomol. 37, 102–124.
6. 岸田泰則(2011)『日本産蛾類標準図鑑II』学研教育出版.
7. 川邊透、前畑真実(2022)『昆虫変態図鑑』ポプラ社.
8. Martemyanov, V., Bykov, R., Demenkova, M., et al. (2019) Genetic evidence of broad spreading of Lymantria dispar in the West Siberian Plain. PLoS ONE 14(8), e0220954.
9. 長谷川政美(2020)『共生微生物からみた新しい進化学』海鳴社.
10. Higashiura, Y., Yamaguchi, H., Ishihara, M., et al. (2011) Male death resulting from hybridization between subspecies of the gypsy moth, Lymantria dispar. Heredity 106, 603–613.
11. Ilinsky, Y.Y., Tokarev, Y.S., R.A. Bykov, R.A., et al. (2017) Detection of bacterial symbionts (Wolbachia, Spiroplasma) and eukaryotic pathogen (Microsporidia) in Japanese populations of gypsy moth species (Lymantria spp.).
12. 海野和男(2022)『ダマして生きのびる虫の擬態』草思社.
13. Ratcliffe, J.M., Fullard, J.H. (2005) The adaptive function of tiger moth clicks against echolocating bats: an experimental and synthetic approach. J. Exp. Biol. 208, 4689-4698.
14. Barber, J.R., Conner, W.E. (2007) Acoustic mimicry in a predator–prey interaction. Proc. Natl. Acad. Sci. USA 104, 9331–9334.
15. Cheng, T., Wu, J., Wu, Y., et al. (2019) Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nature Ecol. Evol. 1, 1747–1756.
*もっと「進化」について知りたい人の入門書:
☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。





{kind=link}