
はるかむかし地球上のあらゆる生物の共通の祖先がいた。
まだ見ぬその姿は細菌のようだったのだろうか?
ダーウィンが夢見た、共通祖先から連なる全生物の進化の系統樹が、
生物のもつDNA情報にもとづいて、いまその姿を現しつつある。
進化生物学者である著者が撮影した貴重な写真コレクションを用いて、
動物界、なかでも昆虫綱、哺乳綱、鳥綱の驚きの進化を100点超の系統樹で表現する。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。最新刊は『ウイルスとは何か』(仮題、中公新書、2023年1月刊行予定)。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

すべての生き物をめぐる
100の系統樹
第1話
全生物界の系統樹マンダラ
文と写真 長谷川政美
◎「系統樹マンダラ」とは何か
地球上に生息する生物は細菌を除いても870万種に達するという試算があるが、細菌も含めあらゆる生物は一つの共通祖先から進化した。
チャールズ・ダーウィンもあらゆる生物は一つの共通祖先に由来すると考えたが、このことは20世紀後半になって、バクテリアからヒトに至るまであらゆる生物で、細胞の基本的な仕組みが共通であることが明らかになったことで、確かめられた。共通祖先から枝分かれを繰り返しながら多様な種が進化したのである。
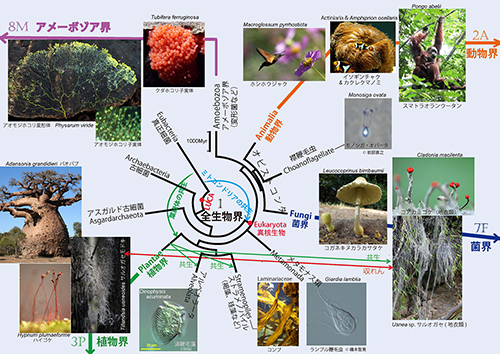
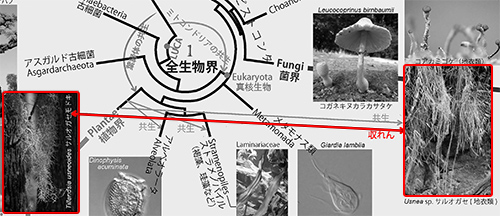
このような進化の様子を表現するのが「系統樹」である。近年、生物のもつDNAの情報から系統樹を構築する分子系統学が発展し、全生物界の進化の概要が明らかになってきた(1)。
系統樹には、さまざまな表わし方があるが、仮想的な(存在したことは確かであるが、どのようなものかはっきりしないので、「仮想的」とした)共通祖先を中心に配置して、そこから放射状に生物が種分化しながら進化してきた様子を表現する「系統樹マンダラ」という方法がある。
そのような表現法の長所は、枝分かれの順番を図の中心部で表現すれば、まわりの広いスペースにたくさんの生物の画像を張り付けることができることにある。一つの共通祖先から出発して多様な生物が進化してきた様子を表現するには、このような方法が視覚的に分かりやすいであろう。
密教の「マンダラ(Mandala)」はたくさんの尊像がある法則に従って配置されたものであり、この世界を表現している。系統樹マンダラは進化の歴史に従って生物の画像が配置され、全生物界の成り立ちを表現する。
Mandaには、サンスクリット語で「中心」あるいは「円」という意味がある。中心に位置する共通祖先から多様な生物が進化してきた様子を表現するのが、系統樹マンダラである。
ダーウィンは1857年9月26日にトマス・ハックスレーに宛てた手紙で、「私自身が生きているうちに見ることはかなわないと思いますが、いつの日かそれぞれの生物界について真の系譜の樹が解明される時代が到来することでしょう」と書いている(2)。
20世紀の後半から急速に発展した分子系統学の成果として、全生物界の系統樹が描かれるようになってきた。まさにダーウィンの夢が実現されつつある時代が到来したのである。細かい点に関しては、今後とも改訂が繰り返されるであろうが、基本的な部分はかなりはっきりと解明されてきたと考えられる。
今回の連載では、毎週1点ずつで、合計およそ100点の系統樹マンダラを用いて、全生物界の進化の様子を視覚的にとらえることを試みる予定である。
京都の東寺にある国宝の「両界曼荼羅」には、胎蔵界と金剛界の2つがある。例えば「胎蔵曼荼羅」には、決して大きいとはいえない183.6×163.0cmのなかに合計420の尊像が描かれている。従ってそれら個々の尊像は非常に小さいが、その細部も詳しく描かれている。このことは、石元康博のB4変形版写真集の各ページいっぱいに印刷された個々の尊像の写真を見るとよく分かる(3)。
「神は細部に宿りたもう」という言葉があるが、マンダラでは全体の構成とともに細部も重要なのである。今回の連載では、系統樹マンダラの図版は拡大できるようにして、なるべく細部まで確認できるようにする予定である。
◎あらゆる生物の共通祖先LUCA
地球はおよそ46億年前に誕生した。その頃の地球は、生命のない星だったが、40億~25億年前の始生代の時期に最初の生命が生まれたと考えられている。最近、日本の探査機「はやぶさ2」が小惑星リュウグウから持ち帰った石や砂からアミノ酸が23種類見つかったことが発表され、話題になっている(5)。最初の生命のもとになったアミノ酸が、隕石として地球に衝突した小惑星からもたらされた可能性があるのだ。生命のもとになった物質は、最初は無生物的に作られたと考えられる。
最初の生命がどのようにして生まれたかについては、現在でもよく分かっていないが、地球上の多様な生き物が一つの共通祖先から進化してきたことは確かであろう。動物、植物、菌類(真菌)、さらに細菌に至るまで、細胞や生命の基本的な仕組みが共通であることから、それぞれが独立に生まれたとは考えられないのである。
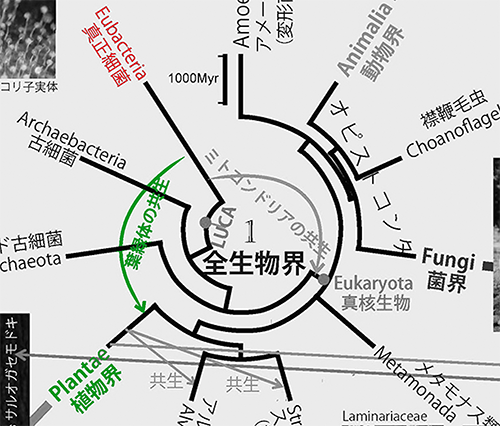
動物、植物、菌類、細菌などすべての生き物の祖先になったものを「LUCA(the Last Universal Common Ancestor;すべての生物の最後の共通祖先)」と呼ぶ。図1は、LUCAから「真正細菌」、「古細菌」、「真核生物」が生まれ、真核生物のなかから動物、植物、菌類、アメーボゾア、そのほかさまざまな生物が生まれたことを表わしている(下図は図1の中央部分を拡大して「LUCA」を彩色)。
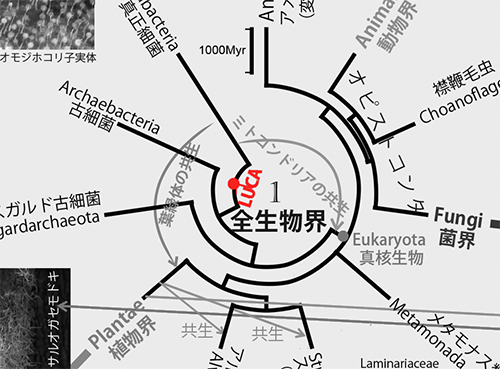
このようにあらゆる生物は、細胞の構造からは原核生物と真核生物に分けられるが、1970年代後半から原核生物のなかに大きく異なる2つの系統があることが明らかになってきた。
従来から知られていた大腸菌やシアノバクテリアなどの真正細菌とそれらとは違うメタン細菌やハロバクテリアなどの「古細菌」である。ところが真正細菌、古細菌、それに真核生物三者のあいだの系統的な関係が不明であった。
1989年になってはじめてこの問題が解決し、古細菌が真正細菌よりも真核生物に近縁であることが明らかになったのだ(6)。その後、それまで古細菌と見なされてきたアスガルド古細菌がほかの古細菌よりもさらに真核生物に近縁であることが判明した(7)。従って、図1で示すように、真核生物はアスガルド古細菌と共通の祖先から進化したことになる(下図は図1の中央の該当部分を拡大して彩色)。
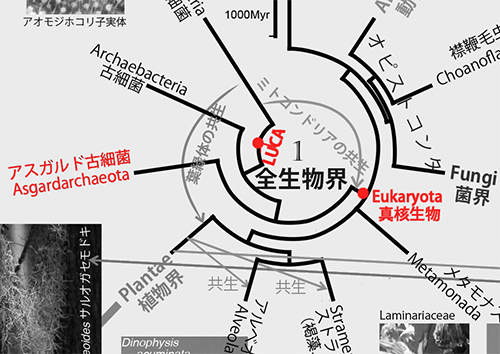
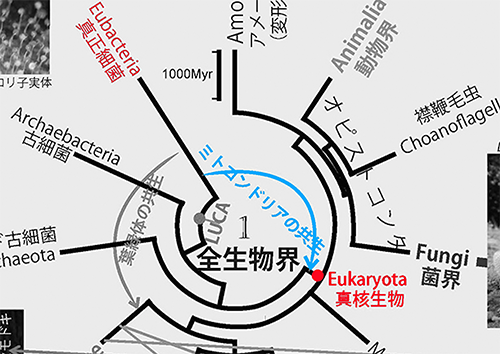
ミトコンドリアは真核生物の細胞の中に存在し、たいていの真核生物における好気呼吸の場である。ミトコンドリアには細胞核にあるゲノムとは別に独自のゲノムがあり、分子系統学的な解析からも真正細菌のなかのアルファプロテオバクテリアが共生してできたものであることが明らかである。
アスガルド古細菌に近縁な真核生物の祖先細胞内にアルファプロテオバクテリアが共生し、それがミトコンドリアに進化したのである。ミトコンドリアがなければ、われわれ動物における高度な運動能力は実現できなかったであろう。
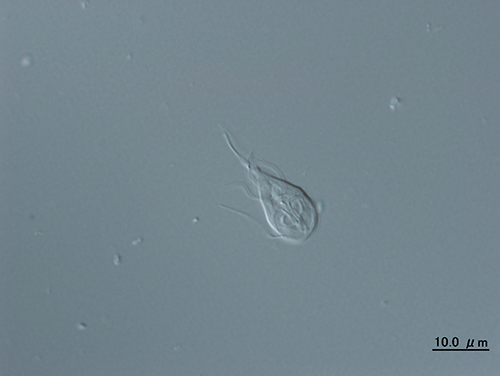
ランブル鞭毛虫はヒトにも感染し、下痢などの症状を起す寄生虫であるが、ミトコンドリアをもたない。当初そのような生物はミトコンドリアが共生する以前の原始的な真核生物の系統だと考えられたが、1990年代後半になるとそのような真核生物でもかつてミトコンドリアをもっていた痕跡が見つかるようになってきた。
従って、ランブル鞭毛虫は、寄生生活に適応した結果、ミトコンドリアを失ったものと考えられる(8)。ミトコンドリアをもたない真核生物はいろいろあるが、調べられた限りすべてのものでかつてミトコンドリアをもっていた痕跡が見つかったのである。
今回の連載は、生き物の写真を使った系統樹マンダラが主体なので、真正細菌や古細菌などの原核生物にはあまり触れない。形態的には彼らの多様性は限られているが、彼らは真核生物にくらべてはるかに多様な代謝システムを進化させており、共生細菌叢を通じてわれわれヒトもその恩恵を受けており、われわれは細菌によって生かされているという見方も可能である(4)。
◎真核生物の多様化
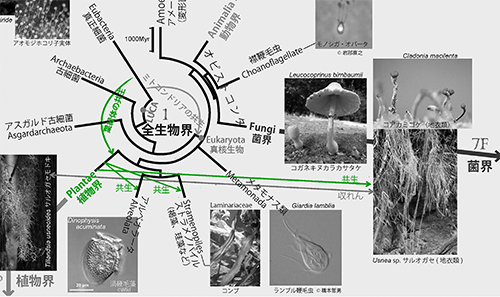
ミトコンドリアの共生と似たようなことが、葉緑体の起源にも見られる。植物がもつ葉緑体は、真正細菌のシアノバクテリアが植物の祖先細胞に共生してできたものである(図1の緑色の矢印)。
光合成する真核生物には、植物以外にも渦鞭毛藻、褐藻、珪藻などがあるが、これらは葉緑体をもった緑藻がそれぞれの祖先細胞内に共生し、光合成ができるようになったものである。また地衣類は菌類と藻類の共生体である。図1ではこれらの共生も緑色の矢印で示した。このように共生は生物進化のさまざまな場面で重要な役割を果たしてきた(下図は図1の該当部分を彩色)。
たくさんのばらばらな系統を含む原生生物の系統関係は複雑だが、動物界、植物界、菌界のあいだの関係はどうなのだろうか。1980年代、私はそのような疑問を抱いていた。
これは、生物のかたちや生き方などを見ていてもまったく手掛かりが得られない問題である。ところが、DNAには確かな手掛かりがあるのだ。あらゆる生物は細胞内にたんぱく質を合成する工場であるリボソームをもつ。リボソームはたくさんのたんぱく質とリボソームRNA と呼ばれる2種類の大きなRNAで構成されている。あらゆる生物は、リボソームRNAをもつから、それを用いてこの問題に挑戦することができた。
その頃蓄積しつつあったリボソームRNAの塩基配列データを解析してみた結果、動物界と菌界が植物界よりも近縁になった。リボソームRNAには、18Sと28Sという2種類のものがあるが、どちらを使っても、動物と菌類が近縁という結果になったのである(9)。
現在から見ると、結果的には正しいものであったが、当時の段階では系統樹推定の誤差も大きく、確定的なものではなかった。ところがその後、1990年代になると、次第にこのような関係が受け入れられるようになっていった。
こうして、菌類は動物や次に述べる襟鞭毛虫などとともに、オピストコンタ(Opisthokonta/後方鞭毛生物ともいう)というもっと大きな分類単位に入れられることになった。オピストコンタの姉妹群(いちばん近縁なグループ)が変形菌などのアメーボゾアであり、植物はもっと遠い関係にあることが明らかになった(下図は図1の該当部分を彩色)。
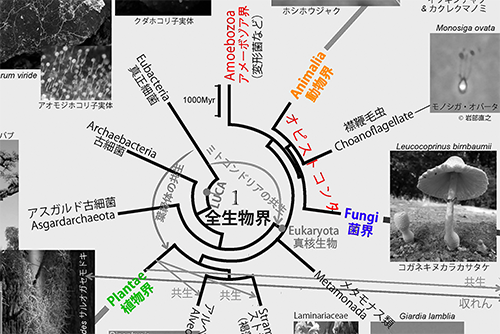
襟鞭毛虫は細菌を捕食して生きている。襟鞭毛虫はロゼット状(タンポポの葉が地中から直接出て放射状に並んでいる様子)のコロニーを作るが、それは多細胞動物の初期段階を彷彿とさせるものである。襟鞭毛虫と動物の共通祖先が生きていたのはおよそ10億年前であったが、その頃から動物の進化が始まったと考えられる(4)。
サルオガセという地衣類(子囊菌という菌類に藻類が共生したもの)があるが、見かけ上これによく似たサルオガセモドキ(単子葉植物イネ目パイナップル科)という植物がある。サルオガセは樹皮に付着して垂れ下がる糸状の地衣類だが、サルオガセモドキもそっくりな形態である。
ただし、この連載で取り上げる予定の分類群は、私自身の写真のコレクションがある程度そろっているものについてだけであり、動物界、なかでも昆虫綱、哺乳綱、鳥綱に偏ったものである。あくまでも、あらゆる生物を含めた系統樹マンダラ構築の第一歩である。これ以外のものについては、将来このような「全生物界系統樹マンダラ」をもっと充実させようとする人々が現れることを期待している。
この連載で予定している100点ほどの系統樹マンダラはそれぞれ独立したものではなく、それぞれの中心に位置する祖先は、元をたどれば図1のLUCAに由来するものである。あらゆる生物が、このようなかたちでお互いに結びついていることを実感して頂ければ幸いである。
ここで分類単位について説明しておこう。分類単位には大きな順から「超生物界(Domain/ドメイン)」、「界(Kingdom)」、 「門(Phylum)」、「綱 (Class)」、「目(Order)」、「科(Family)」、「属(Genus)」、「種(Species)」などがある。
ドメインとは真正細菌、古細菌、真核生物などに対応するが、古細菌のなかでアスガルド古細菌はほかの古細菌よりも真核生物に近縁なので、アスガルド古細菌を古細菌に含めると、古細菌ドメインは成り立たない。界とは動物界、植物界、菌界、アメーボゾア界などである。
分類の対象が増えてくると上に挙げた分類単位だけでは足りなくなり、界の下の分類単位である「亜界(Subkingdom)」、いくつかの目を統合した「上目 (Superorder)」、目の下の分類単位としての「亜目 Suborder」、いくつかの科を統合した「上科(Superfamily)」、科の下の分類単位としての「亜科 (Subfamily)」、いくつかの属を統合した「族 Tribe」などが用いられる。
このような階層的な分類が成り立つ背景には、共通祖先から順次枝分かれしながら、多様な生物が進化してきたということがある。現生の生物から逆に祖先をたどっていったときに、同じ属、科、目のメンバーの順に共通の祖先が古くなるのである。
【引用文献】
1. 長谷川政美(2020)『進化38億年の偶然と必然-生命の多様性はどのようにして生まれたか』国書刊行会.
2. Darwin, F., Seward, A.C. eds. (1903) “More letters of Charles Darwin. A record of his work in a series of hitherto unpublished letters”. Vol. 2, p. 104. John Murray.
3. 石元康博(2011)『両界曼荼羅』平凡社.
4. 長谷川政美(2020)『共生微生物からみた新しい進化学』海鳴社.
5. Nakamura, E., Kobayashi, K., Tanaka, R., et al. (2022) On the origin and evolution of the asteroid Ryugu: A comprehensive geochemical perspective. Proc. Jpn. Acad., Ser. B 98, 227-282.
6. Iwabe, N., Kuma, K., Hasegawa, M., Osawa, S., Miyata, T. (1989) Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proc. Natl. Acad. Sci. USA, 86:9355–9359.
7. Eme, L., Spang, A., Lombard, J., et al. (2017) Archaea and the origin of eukaryotes. Nature Rev. Microbiol. 15, 711-723.
8. Hashimoto, T., Sanchez, L., Shirakura, T., Müller, M., Hasegawa, M. (1998) Secondary absence of mitochondria in Giardia lamblia and Trichomonas vaginalis revealed by valyl-tRNA synthetase phylogeny. Proc. Natl. Acad. Sci. USA, 95:6860–6865.
9. Hasegawa, M., Iida, Y., Yano, T., Takaiwa, F., Iwabuchi, M. (1985) Phylogenetic relationships among eukaryotic kingdoms inferred from ribosomal RNA sequences. J. Mol. Evol., 22:32–38.
*もっと「進化」について知りたい人の入門書:
☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。





{kind=link}