
はるかむかし地球上のあらゆる生物の共通の祖先がいた。
まだ見ぬその姿は細菌のようだったのだろうか?
ダーウィンが夢見た、共通祖先から連なる全生物の進化の系統樹が、
生物のもつDNA情報にもとづいて、いまその姿を現しつつある。
進化生物学者である著者が撮影した貴重な写真コレクションを用いて、
動物界、なかでも昆虫綱、哺乳綱、鳥綱の驚きの進化を100点超の系統樹で表現する。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。最新刊は『ウイルスとは何か』(仮題、中公新書、2023年1月刊行予定)。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

すべての生き物をめぐる
100の系統樹
第4話
単子葉植物の系統樹マンダラ
文と写真 長谷川政美
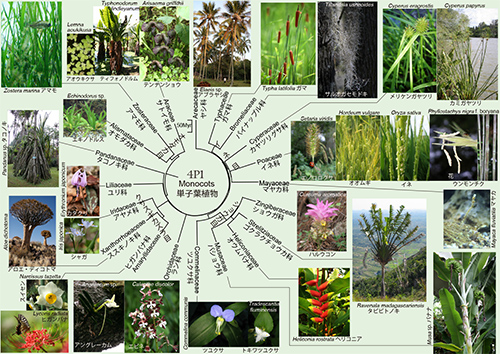
◎イネ科の進化
イネ科はイネ、オオムギ、コムギ、トウモロコシなどわれわれの食料として重要な位置を占めるものであるが、以前はイネ科の主要な系統であるイネやタケなどの系統、コムギやオオムギの系統、トウモロコシやエノコログサの系統という三大系統は、非鳥恐竜が絶滅した6600万年前以降の新生代に生まれたと考えられていた。
ところが草食恐竜の糞化石を分析してみると、恐竜の時代にこの3大系統がすでに分かれていたことが分かってきた(3)。このことは植物食恐竜の糞化石を解析した結果明らかになったことである。

イネ科植物の化石は乏しいので、三大系統がいつ頃分かれたかについての化石からの証拠がほとんどなかったが、植物食恐竜の糞のなかの植物珪酸体がその手掛かりを与えてくれたのである。
植物珪酸体は植物の種類によってかたちが異なる上に、化石として保存される。それを解析した結果、イネ科の三大系統は6600万年前以前の白亜紀の間に揃っていたことが明らかになった。

◎さまざまな方向への進化
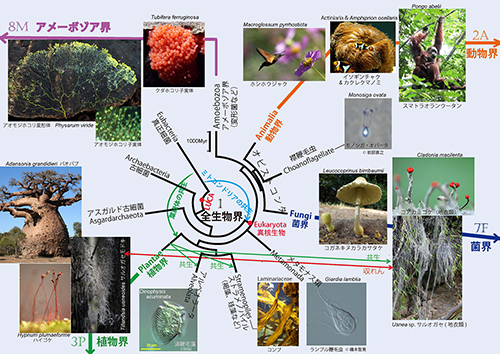
パイナップル科のサルオガセモドキは、下の図1.全生物界系統樹マンダラで菌界の地衣類であるサルオガセとのあいだの収れん進化の例として紹介したものである。
サルオガセモドキは高い木の枝から垂れ下がっているが、木に寄生しているわけではない。この植物は普通の根を失っているが、葉が根の役割を果たしているのだ。葉は細かい毛で覆われ、わずかな雨や露を葉の表面の毛、鱗片、表皮細胞から吸収できるようになっていて、自分で光合成も行って、花も咲かせる。木の表面を流れ落ちる雨水やほこりなどが養分の供給源になっているという(4)。
このような水生適応した植物の進化は、水から陸上に進出した四足動物(陸上脊椎動物)のなかでも、哺乳類に限ってもクジラ、ジュゴン、アザラシなどたくさんの系統が独立に水生に戻ったことと似ている。
最初に陸上植物や四足動物が陸上に進出した頃には、そこは競争相手がいない新天地だったが、次第に空いている生態的地位(ニッチェ)がなくなってきて、水生に戻るものが現れたのである。

図にあるテンナンショウはブータンのもので、「ゾウの耳」とも呼ばれる。このゾウの耳のように見えるのは、花の集まりを包む苞葉というものであり、ミズバショウにも同じような構造がある。
◎植物と動物の共進化
ラン科は従来クサスギカズラ目Asparagalesに入れられていたが、分子系統学からはこれに入らない可能性が指摘されている(2;その後のさらに大規模な系統解析の結果、再びラン科もクサスギカズラ目に戻されたようである:長谷部光泰氏私信)。
ラン科は単子葉植物のなかで最も種数の多い科である。この科はいくつかの亜科から成るが、最大の亜科が15,000種以上を含むセッコク亜科 Epidendroideaeであり、図4P1のなかのマダガスカルのアングレーカム属Angraecumがその一部である(6)。
セッコク亜科の大部分は熱帯の着生植物であり、下の写真は鳥の糞に含まれた種子が木の枝に着生したものである。

実はこれと同属のAngraecum sesquipedaleというランは、もっと長い距をもっている。19世紀に園芸植物としてイギリスに入ってきたこのランを見たチャールズ・ダーウィンは、マダガスカルにこんなに長い距をもったランがあるからには、そこにはこの距の奥まで届くような長い口吻をもったガがいるに違いないと予言した。
ダーウィンの死後1903年になって、彼が予言した通りのスズメガが発見され、Xanthopan morganii praedictaと命名された。最初はアフリカにいるキサントパンスズメガXanthopan morganiiの亜種とされたのだが、亜種名のpraedictaは「予言されたもの」という意味である。最近は、マダガスカルの「予言されたもの」をXanthopan praedictaと独立種にする論文も増えている。
植物には花粉を昆虫などの動物に運んでもらって受粉を助けてもらうものが多い。受粉を助ける昆虫のほう(送粉者という)は、蜜や花粉などを食料としてもらうので、双方にとって利益がある。その際植物にとっては、ほかの種類の植物の花粉を運んでこられたのでは困る。なるべく、自分と同じ種の花粉だけを運んでもらうのが望ましいのだ。一方、昆虫の側からは、自分だけが蜜にありつけることが望ましい。
このように、双方の利益が合致した方向に進化が進んだ結果、現在のAngraecum sesquipedaleとXanthopan praedictaが進化したと考えられる。このような進化を「共進化」という。こうしてAngraecum sesquipedaleの長い距の奥にたまった蜜を吸えるのは、長い口吻をもったXanthopan praedictaだけということになったわけである。

またマダガスカルには、Xanthopan praedictaほどの長さではないが、さまざまな長さの口吻をもったスズメガもいる。植物と送粉者の間の共進化によって、それぞれ距と口吻の長さが決まってきたのであろう。
アングレーカム属のランと送粉者であるガの系統樹解析の結果、Angraecum sesquipedaleが姉妹群(一番近縁なもの)から分かれたのがおよそ750万年前であり、一方送粉者のXanthopan praedictaがアフリカの姉妹群(Xanthopan morganii)から分かれたのもほとんど同じ頃だったと推定される。
つまり、アフリカからマダガスカルに渡ってきたこのスズメガの祖先が新たなパートナーとしてアングレーカム属のランを選び、その後の両者のあいだの共進化の結果、今日見られるようなものが成立したと考えられる(7)。
つづく
【引用文献】
1. Angiosperm Phylogeny Group (2016) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linnean Soc. 181, 1–20.
2. Hertweck, K.L., Kinney, M.S., Stuart, S.A., et al. (2015) Phylogenetics, divergence times and diversification from three genomic partitions in monocots. Bot. J. Linnean Soc. 178, 375–393.
3. 長谷川政美、米澤隆弘 (2010) イネ科植物の進化と動物との関わり.科学 80(2), 128–132.
4. 湯浅浩史(2012)『世界の葉と根の不思議-環境に適した進化のかたち』誠文堂新光社.
5. Cusimano, N., Bogner, J., Mayo, S.J., et al. (2011) Relationships within the Araceae: Comparison of morphological patterns with molecular phylogenies. Amer. J. Bot. 98(4), 654-668.
6. Freudenstein, J.V., Chase, M.W. (2015) Phylogenetic relationships in Epidendroideae (Orchidaceae), one of the great flowering plant radiations: progressive specialization and diversification. Ann. Bot. 115, 665–681.
7. Netz, C., Renner, S.S. (2017) Long-spurred Angraecum orchids and long-tongued sphingid moths on Madagascar: a time frame for Darwin’s predicted Xanthopan/Angraecum coevolution. Biol. J. Linnean Soc. 122, 469–478.
*もっと「進化」について知りたい人の入門書:
☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。





{kind=link}