
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第34話
細胞核の起源
文と写真 長谷川政美
◎2つの超生物界
真核生物がアスガード古細菌群から進化したものならば、いわゆる古細菌は系統的にまとまったグループではないことになる。そのために、これまでの「真正細菌」「古細菌」「真核生物」の3つの超生物界への分類に代わって、古細菌と真核生物をあわせた「アーケアArchaea」という超生物界を設け、これと「真正細菌Bacteria超生物界」の2つの分類単位に地球上のあらゆる生物を分類するべきかもしれない(図34-1)。
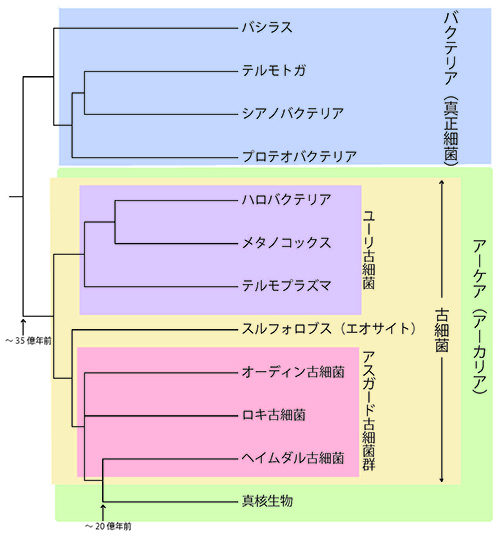
図34-1 地球上の生物の2大分類群:「バクテリア(真正細菌)」と「アーケア」。アーケアは古細菌と真核生物を含む超生物界として定義しなおされたもの。古細菌Archaebacteriaと核をもった生物Karyotaをあわせた「アーカリアArkarya」という分類名も提案されている。この流れは、ヘイムダル古細菌などのアスガード古細菌群がほかの古細菌よりも真核生物に近縁であることが明らかになって加速されたが、ジム・レイク(図32-1)が1980年代から主張してきたエオサイト説に起源がある。レイクはリボソームの形態などからスルフォロブスなどエオサイトと呼ばれる古細菌が、ハロバクテリアやメタノコッカスなどユーリ古細菌と呼ばれるほかの古細菌よりも真核生物に近いと考えたのである。彼はエオサイト+真核生物を「Karyota」と呼んだ。
1989年に岩部・宮田らのグループが重複遺伝子の複合系統樹解析によって古細菌が同じ原核生物の真正細菌よりも真核生物に近縁であることを示した(第30話)。これを受けて、1990年にカール・ウースは「真正細菌Eubacteria」「古細菌Archaebacteria」「真核生物Eukaryotes」に代わって、「バクテリアBacteria」「アーケアArchaea」「ユーカリアEukarya」という分類名を使うことを提案した。
しかしながら同じ細菌類の真正細菌だけを「Bacteria」と呼び、もともと自分が命名した「古細菌Archaebacteria」をあたかも細菌ではないかのような呼び方をすることには批判があった。20世紀の前半に進化の総合学説の建設に貢献したエルンスト・マイヤーもそのような批判者の一人であった(図34-2)。
図34-2 エルンスト・マイヤーErnst W. Mayr(1904-2005)。1987年、カリフォルニア大学バークレー校での講演会にて。
古細菌が単系統のグループだとすると、それが真核生物に近いというだけの理由で、勝手に名前を変えることには納得がいかなかったのである。しかし、ウースの権威に押し切られるかたちで、多くの研究者が新しい名前を使うようになった。その頃は私自身も古細菌は単系統のグループだと考えていたので、アーケアという呼び方には抵抗を感じて、長いあいだこれを使ってこなかった。
2010年、Nature誌に米澤隆弘君と共著で簡単なコメント論文を書いた際に、編集者から「Archaebacteria」を「Archaea」に直すように求められたが、そのような理由を書いて断ったということもあった。しかし、いわゆる古細菌が側系統のグループであることが明らかになった現在、古細菌と真核生物をあわせた超生物界の名前として「アーケアArchaea」を使うのは妥当かもしれないと考えている(図34-1)。
◎イントロンと細胞核の起源
真核生物が、真正細菌や古細菌などほかの生物と違う最大の特徴は、その名前にもある「核」をもっていることにある。核は核膜に囲まれたものであり、そのなかにDNAはクロマチンというかたちで存在する。核は真核生物固有のものであり、このような構造がどのように進化したかは、謎につつまれていた。
1970年代末の分子生物学の思いがけない発見に第22話で出てきた「イントロン」がある。真核生物のたんぱく質をコードしている遺伝子の多くで、アミノ酸をコードしているDNAの塩基配列がアミノ酸のコードとは関係ない塩基配列で分断されているという発見である(図34-3)
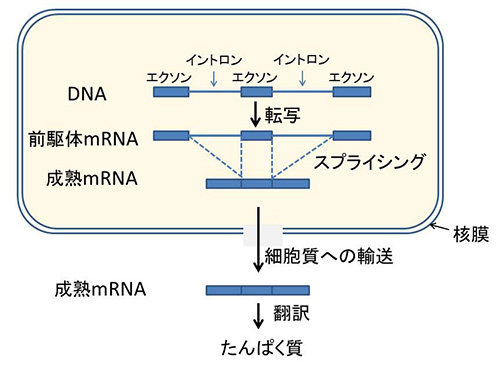
図34-3 イントロンのスプライシング。マーチンとクーニンは、mRNAのスプライシングが終了するまで、mRNAをリボソームから隔離しておくために核膜が進化したと考えた。
アミノ酸をコードしている領域は「エクソン」、途中に介在している領域は「イントロン」と呼ばれている。イントロンの数は遺伝子によって異なるが、10を超えることもあり、しかも一つひとつのイントロンは長く、エクソンよりも長いことが多い。
遺伝子が発現する際には、DNAからまずイントロンも含めた全領域が転写されて、前駆体メッセンジャーRNA(前駆体mRNA)が合成される。続いてイントロンの部分が除去され、隣り合ったエクソンが連結されるが、この過程を「スプライシング」という。こうして、成熟メッセンジャーRNAができるが、ここまでの過程は細胞核のなかで進行する。続いて成熟メッセンジャーRNAは細胞質に輸送され、この分子のもつ情報に従ってリボソームでたんぱく質が合成される。
この一見無駄に思われるイントロンの起源をめぐっては、発見後30年ほどのあいだ、激しい論争が続いた。すべての生き物の最後の共通祖先であるLUCAは、イントロンをもっていたが、真正細菌や古細菌ではその後の進化で失われたという説(イントロン前生説)と、イントロンは真核生物のゲノム中に後から挿入されたという説(イントロン後生説)である。
しかし現在ではイントロン後生説が有力である。真核生物と共通の遺伝子で、真正細菌や古細菌ではイントロンが見つからないことをすべてイントロンの消失で説明するのは難しいということと、後で挿入されたと思われる真核生物のイントロンが存在するという理由からである。
前に重複遺伝子の話をしたが、遺伝子がゲノム中で重複を繰り返して、数十個の遺伝子から成る遺伝子ファミリーができることがある。通常、それぞれの遺伝子は少しずつ異なる機能に特化することにより、遺伝子ファミリー全体で多様な機能を果たせるようになる。
ビル・マーチン(図32-2)とユージン・クーニンは、イントロンの侵入が真核生物誕生の頃に集中的に起ったのではないかと考えた。そのことを示すために彼らは、遺伝子ファミリーを「太古のパラログ」と「新しいパラログ」の2つに分類して、イントロンの位置を調べた。
◎マーチンとクーニンの仮説
遺伝子が重複したあとで異なる種に分かれた場合、同じ重複遺伝子の異なる種間の関係を「オーソログ」、異なる重複遺伝子間の関係を「パラログ」という(図34-4)。
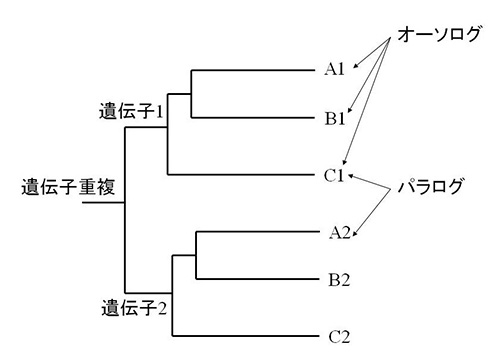
図34-4 オーソログとパラログ。
太古のパラログとは、すべての真核生物に見られるが、原核生物では重複のない遺伝子ファミリーと定義される。従って、太古のパラログを生み出した遺伝子重複は、真核生物の最後の共通祖先よりも以前の、真核生物が初期進化を遂げた時代に起ったものと考えられる。
一方、新しいパラログは、真核生物の一部のグループ、例えば動物界だけでしか見られない遺伝子ファミリーである。この場合は、遺伝子重複はその特定のグループが進化した途中で起ったものと推定される。
マーチンとクーニンは、真核生物が誕生した時期に集中的にイントロンが侵入したのならば、さまざまな遺伝子にランダムに入ったはずだと考えた。従って、その頃に遺伝子重複を繰り返していた太古のパラログでは、遺伝子ファミリーのメンバーによってイントロンの位置が異なることが予想される。
一方、新しいパラログでは、遺伝子重複の前にイントロンの侵入があったと考えられるので、イントロンの位置が保存されていることが予想される。実際に彼らが調べてみたところ、まさに予想した通りであった。太古のパラログのイントロンの位置はさまざまであるが、新しいパラログではイントロンの位置が保存されていたのである。
マーチンとクーニンは、この結果を土台にして、細胞核の起源までをも説明する壮大な仮説を提唱した。水素仮説が文字通り正しいかどうかは分からないが、プロテオバクテリアが古細菌に内部共生し、ミトコンドリアやヒドロゲノソームに進化したのは確かであろう。そのような内部共生の初期実験で、内部共生体となったプロテオバクテリアは宿主細胞内に複数個存在したであろう。その際、死んだ内部共生体は溶解し、そのDNAは宿主の細胞質に流れ込んで、宿主細胞のゲノムに組み込まれたであろう。そのなかには、たんぱく質をコードする遺伝子がそっくり組み込まれて、宿主の側でたんぱく質が合成されるようになったものもあるだろう。
現在、ミトコンドリアを作っているたんぱく質の遺伝子の多くは核ゲノムにあるが、このような共生体から宿主への遺伝子転移がうまく働くためには、遺伝子産物が共生体に運ばれるようなシステムの進化が必要である。転移した遺伝子のなかには、遺伝子産物がミトコンドリアに輸送されず、細胞質のなかで働くようになったものもあるだろう。イントロンの多くは、このように溶解した内部共生体のDNA断片が宿主のゲノムに組み込まれるときに、ランダムに入り込んだものと考えられる。
古細菌や真正細菌などの核をもたない原核生物では、たんぱく質をコードする遺伝子の転写と翻訳は共役している。つまり、遺伝子がメッセンジャーRNAに転写されるにつれて、そのRNAにたんぱく質合成装置であるリボソームが付着して、たんぱく質の合成を開始する。しかし、イントロンを含む遺伝子をもつ真核生物でそのようなことが起ったら、イントロンの部分もいっしょに翻訳されてしまうことになる。
マーチンとクーニンは、たんぱく質合成の場であるリボソームをゲノムと隔離するために核膜を進化させる必要があったのではないかと考えたのだ。イントロンを含む遺伝子からたんぱく質を合成する前に、前駆体メッセンジャーRNAをスプライシングして成熟メッセンジャーRNAにしなければならない。しかし、スプライシングが完了する前に、たんぱく質の合成が始まってしまったら、全く役に立たないたんぱく質ができてしまうことになる。これを避けるために、ゲノムをリボソームのある細胞質から隔離するための核膜を進化させ、核のなかでスプライシングを完了させてから、成熟メッセンジャーRNAをリボソームに届けるようなシステムが必要だったというのが、マーチンとクーニンの説の要点である(図34-3)。
ゲノムとリボソームをなるべく隔離できるような膜ができれば、スプライシングが完了する前にたんぱく質の合成が始まってしまうという無駄な混乱が減るので、それだけその細胞の適応度が増したと考えられる。
真核生物がもつ特徴のうちで、ミトコンドリアや葉緑体の起源については、細胞内共生説で一応納得のいく説明がなされてきた。ところが、すべての真核細胞がもつ細胞核がどのようにして生まれてきたかという問題は、まったく闇に包まれていた。マーチンとクーニンの「対イントロン説」は、これを説明することを目指した最初の仮説と言える。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


