
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第20話
発生と進化
<繰り返し要素の個性化と多様な形態の進化>
文と写真 長谷川政美
ホメオボックスの発見は、主に2つの点でわれわれの自己認識を拡張することになった。第1に、マスター制御遺伝子は、卵から成体にいたる生物の発生を制御する包括的な遺伝的設計図の存在を示したという点である。第2に、すべての主要な多細胞動物類にかんする研究によって、あらゆる動物類は、組み合わせの似た類縁関係にあるマスター制御遺伝子群をもっていることが明らかになったという点である。このことは驚くべき事実を示している。1億年あるいはもっと長期間にわたる進化において、今日われわれの知るさまざまな生物はすべて単一の発生制御システムによってつくり出されたのである。ハエ、カタツムリ、ヒトデ、魚類、そしてわれわれ自身さえも!
フランク・ラドル (1999)(ワルター・ゲーリング
『ホメオボックス・ストーリー』の序文、浅島誠ら訳)
◎モジュール:繰り返し構造
5億年以上前のカンブリア紀に最初に出現したさまざまな動物に、「モジュール」と呼ばれる繰り返し構造が認められる(図20-1a、b)。


似たような多数の部品が繰り返されているのだ。このような構造は、その後に生まれた恐竜(図20-2a)や哺乳類(図20-2b)にも同じように認められる。節足動物や環形動物の体節や脊椎動物の脊椎などは、これらの門のボディープラン(からだの設計図)の基本単位である。


動物のからだのほかの器官の多くも繰り返し構造でできている。ヒトの歯、手、足やチョウの翅の模様(図20-3)などもそのような例である。繰り返し構造とそれぞれのモジュールの多様化によって複雑なからだができ上っている。

ヒトの歯は、門歯、犬歯、臼歯などに分化している(異歯性Heterodonty)が、哺乳類の共通祖先では同じような歯が並んでいた(同歯性Homodonty)と考えられる。異歯性はモジュール要素が個性化したことによって進化した。繰り返し構造の数が変わるだけではなく、このようにモジュール要素の個性化によって、動物の形態がさまざまに進化することが可能になっている。ホメオティック遺伝子がそこで主要な役割を果たしているのだ。
図20-4に奈良東大寺の大仏の前にある2頭の青銅製のチョウを示した。

◎昆虫の翅の多様化
ハエは双翅目に分類されるが、「双翅」とは翅が2枚しかないことから来ている。昆虫は前翅と後翅あわせて4枚の翅をもっているのが普通である。ところが、ハエなどの双翅目昆虫では後翅が小さく退化して、前翅のような翅脈がなく、先端が丸まって棍棒状になっているのだ。このような後翅は飛翔中のからだの揺れを感知して、バランスをとる役目を果たしているので「平均棍」と呼ばれる(図20-5)。
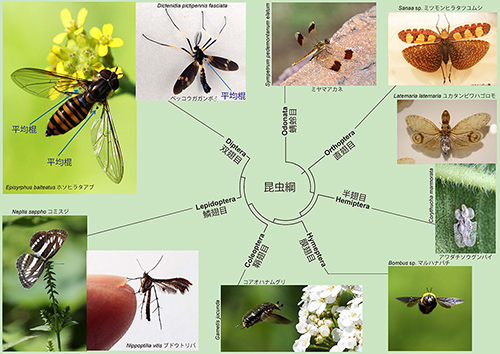
図20-5 昆虫の翅の多様化。昆虫綱のなかで最初にほかから分かれたトンボなどの蜻蛉目では、前翅と後翅はよく似たかたちであるが、そのあとから分かれたグループでは様々に分化している。アブやガガンボなど双翅目昆虫では、後翅が変化して平均棍になっている。コアオハナムグリの写真から分かるように、鞘翅目(甲虫)では、飛翔にはもっぱら膜状の後翅が使われ、硬い鞘状の前翅は飛んでいないときに柔らかい後翅を保護するように機能している。クリックすると大きく表示されます。

図20-6 ヒラタアブ(種名不明:双翅目・ハナアブ科)。
例えば、鞘翅目(甲虫)では、飛翔にはもっぱら膜状の後翅が使われ(図20-5)、硬い鞘状の前翅は飛んでいないときに柔らかい後翅を保護するように機能しているとともに、飛ぶときに体を流線形にする役割もあるという。このような前翅と後翅の分化において、ウルトラバイソラックスのようなホメオティック遺伝子が重要な役割を果たしたのであろう。
図20-5のなかの甲虫、ハナムグリは、花粉を食べに花にやってきているが、このような光景は甲虫の進化のなかでは、比較的最近になってから見られるようになったものである。鞘状の前翅をもった甲虫は、およそ2億6000万年前のペルム紀に出現したと考えられる。その頃にはまだ花を咲かせる植物は出現していなかった。ペルム紀の甲虫は木の幹の隙間に入り込んで、菌類や腐食した植物組織を食べていた。きれいな花を咲かせる被子植物が現れたのはおよそ1億4000万年前の白亜紀前期以降であり、それに伴って花粉を食べる甲虫が進化した。きれいな花は花粉を食べる昆虫を引きつけて受粉を助けてもらうために進化したものであり、植物と動物の共進化の結果である。飛べる昆虫は、遠くまで花粉を運んでくれるので、送粉者としては最適だった。
初期の被子植物の花には蜜がなかったが(被子植物の系統樹で初期にほかから分かれたモクレンの仲間の花には蜜はない)、その後、蜜を提供するものが現われた。昆虫のなかで鱗翅目と呼ばれるガやチョウの仲間の成虫はストローのような口吻をもっており、ハチドリとの収斂進化の例として前に出てきた図7-1の右のホシホウジャクのように、花の蜜だけを吸うように特殊化した。彼らも幼虫の間は噛むことのできる口器で植物の葉を食べている。
鱗翅目の系統樹で最初にほかから分かれたコバネガ科だけは、成虫でも口吻ではない噛むことのできる口器をもっており、それを使って花粉を食べるが、それ以外の鱗翅目の成虫は花の蜜専門でストローのような口吻しかもっていない。このような植物とガやチョウの関係も、共進化の結果である。
◎昆虫の翅の起源
昆虫は節足動物門の甲殻類のなかから進化した。甲殻類付属肢の原型は、二叉型である。多数の節が関節によって連結した外骨格性の肢で、基部から出る2つの枝、つまり内枝と外枝から成るのだ。付属肢の機能に応じて、それぞれの枝の節数が減ったり、1つの枝が消失することもある(つまり二叉でなくなる)。内枝は歩脚(遊泳や歩行に用いる)として、外枝は鰓脚(さいきゃく)として使われることが多い。鰓脚とは水のなかで、効率よく酸素を取り込むための仕組みである。エボデボの研究から、甲殻類の鰓脚から昆虫の翅が進化したことが分かってきた。
4億1600万年~4億4400万年前のシルル紀に、もともと水中で進化したミジンコのような甲殻類のなかから陸上に進出するものが現われ、そのなかから翅を発達させて空中に飛び出すものが現われた。それが昆虫である。その際に、水中で生活するのに重要だった鰓脚を翅に改造したのだ。
ドイツのミカリス・アヴェロフとステファン・コーエンの次のような研究から、そのことが明らかになった。ショウジョウバエの発生の研究から、翅を作るために必要な2種類のたんぱく質が知られていた。その2種類のたんぱく質をコードする遺伝子に欠陥があると、翅が作れないのだ。甲殻類でその2種類のたんぱく質が発現している場所を調べたところ、二叉型の付属肢のうちの外側の枝、つまり鰓脚だけで発現していることが分かったのである。このことから、鰓脚を翅に改造したことにより、昆虫は空に進出できるようになったと考えられる。
このようにからだの前後軸に沿った繰り返し構造のなかでホメオティック遺伝子が異なる働きをすることによって、さまざまなボディープランが作り出されるという基本原理は、節足動物だけでなくわれわれヒトを含む大部分の動物に共通している。
われわれヒトを含めた脊椎動物も体節というブロックでできている。その体節から、脊柱とそれに連結する肋骨や筋肉がセットとなったモジュールができる(図20-2)。発生の過程で、体節は最初みんな同じように見える。しかしやがて、前後軸上の位置に応じて、異なるタイプの椎骨、肋骨、筋肉などが作られていく。これを制御しているのが、節足動物で働いているのと同じホメオティック遺伝子なのだ。
また、脊椎動物の前足と後足の分化も、ハエの前翅と後翅の分化と同じような仕組みで進行することが明らかになった。前足と後足が違った構造に分化できたおかげで、鳥類やコウモリの飛翔能力が進化したのだ(図20-7、図20-8)。図20-2で示した脊椎動物の脊柱の領域によって異なる形態も、ホメオティック遺伝子によって調節されている。

図20-7 空中の一点に留まって(ホバリングしながら)ユスリカを捕食するヒヨドリHypsipetes amaurotis.鳥類のこのようなすぐれた飛翔能力は、歩行用の後足はそのままにして、前足を翼に進化させることで実現できた)。

図20-8 ジャワオオコウモリPteropus vampyrus.哺乳類のコウモリも、鳥類とは違ったやり方で前足を翼に進化させて空に進出した)。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


