
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第4話
共通祖先からの進化
文と写真 長谷川政美
◎最初の「生命の樹」
図4-1は、ジョン・グールドによる鑑定の結果、ダーウィンがガラパゴスから持ち帰ったミソサザイやシメに似た鳥の標本がフィンチであることが判明した頃(1937年)に、ダーウィンが最初にノートに記した枝分かれ図である。
有名な、“I think” (われ思うに)で始まるページでは、「①」と記された共通祖先から枝分かれを繰り返しながら、さまざまな現生種が生み出される過程を表現している。枝の先端に垂直に線が添えられているのが現生種(この図では13種)を表わし、線が添えられてない枝は絶滅した系統である。

図2-2で示したラマルクの枝分かれ図と違って、ダーウィンのものでは現生種はすべて枝の先端に置かれている。ここでダーウィンは共通祖先からの進化をはっきりと意識したのだ。
しかし、その後20年あまり、彼は自分の考えを公にすることはなかった。彼は自分の発見が社会に及ぼす衝撃の大きさを悟ったからである。それでも、1844年に植物学者ジョセフ・フッカー Joseph Hooker に宛てた手紙のなかで、自分の心情を次のように告白している。
私はガラパゴスの生物分布やアメリカの化石哺乳類の形質に大変心を打たれたので、種とはなにか、という問題に関わるものならば何でも集めてみようと決心しました。……そしてついに一条の光が差し込んだのです。その結果、当初の私の考えとはまったく逆に、種は変わり得ないものではないことを(これは殺人を告白するようなものですが)ほぼ確信するに至りました。
ダーウィンが自説の公表をためらったのは、それよりも300年近く前のコペルニクスの置かれた状況とよく似ていた。コペルニクスが地動説の着想を得たのは1508~1510年頃と推定されているが、当初は署名なしの手稿を知人たちに閲覧させるだけで、40年あまりこれを公表することはなかった。彼の主著『回転論』が出版されたのは、彼が亡くなる1543年のことであった。これら科学史上の2大革命は、それだけ社会に及ぼす衝撃も大きかったということであろう。
◎『種の起源』の出版
1859年になって、ダーウィンは『種の起源』を出版するが、彼がようやく自説の公表に踏み切ったきっかけは、前の年に受け取った手紙であった。差出人は、現在のインドネシアにあるモルッカ諸島テルナテ島にいたアルフレッド・ラッセル・ウォーレス Alfred Russel Wallace (1823–1913)であった。手紙にはウォーレス自身の新しい論文の原稿が添えられていた。論文のタイトルは「変種がもとの型から無限に遠ざかる傾向について」。そこには、ダーウィン自身が長年にわたって考えを温めてきた自然選択説と同じ進化の理論が展開されていたのだ。
びっくりしたダーウィンは、このままでは自分の先取権が奪われると考えて、それまでにも個人的に自分の新しい考えを聞いてもらっていた地質学者のチャールズ・ライエルと先ほど出てきたジョセフ・フッカーに相談した。その結果、ウォーレスの論文と一緒にダーウィンの草稿の抜粋が、ロンドンにあるリンネ協会の例会で発表された。それまでにダーウィンは、20年ほどにわたっていずれ刊行されるように、大著の原稿を少しずつ準備していたが、ウォーレスの手紙を契機に方針を変えたのだ。予定していた大著の要約版というかたちで急きょ出版されたのが『種の起源』であった。この本は、通常『種の起源』と呼ばれるが、原著のタイトルは、「自然選択すなわち存続をめぐる争いにおいて有利なものが保存されることによる種の起源」となっている。
『種の起源』には図版がほとんどないが、唯一のものが図4-2である。これは、「自然選択」と題した第4章に現れる。下にある「A」から「L」までの祖先種から枝分かれを繰り返しながら一番上にある現生種が進化してきた様子が表現されている。
そこには現在に至る子孫を残さずに途中で絶滅したたくさんの系統も示されている。例えばAの子孫はたくさんの種に分かれ、たくさんの現生種を残したが(それでもたくさんの絶滅した系統も含む)、それに近いB、C、D、Eの子孫は途中で絶滅したように描かれている。
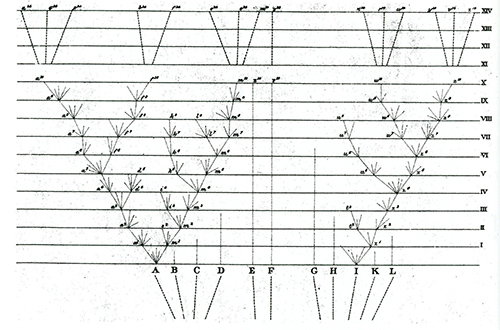
Aから伸びた系統で、 a1や a2から短い線が出ているのは、変異があることを示し、それぞれそのうちの一つの系統だけが生き残る。そこに働く原理が、「自然選択 Natural selection 」である。たくさんの変異のなかで、子孫を残す上で有利な変異が選ばれて残るというのが、自然選択の考えである。Natural selection は「自然淘汰」と翻訳されることもあるが、「淘汰」には「ふるい落す」という意味が強調されており、ここではむしろ「選ばれて残る」ことを強調して「自然選択」としておく。
a3からは a4と d4の2種が枝分かれで生ずる。種分化である。 d4の系統はその後まもなく絶滅してしまうが、 a5から種分化で生まれたf5の系統は現在まで存続する。このように、図4-2は複数の現生種が共通の祖先から進化する様子をはっきりと表現している。
ダーウィン以前は進化論であっても「自然の階段」つまり「存在の偉大な連鎖 Great chain of being」というとらえ方であったが、ダーウィンは「生命の偉大な樹 Great tree of life」という新しい視点を与えたのである。彼は、リンネの階層的分類体系が、生物進化の歴史を表現するものであることを明らかにしたのだ。フンボルトは種の間の生態的なつながりを見出したが、ダーウィンは種の間の遺伝的なつながりを見出したといえる。
ダーウィンは、共通祖先からの進化を、「変化を伴う継承(由来)Descent with modification」と呼んだ。彼は1858年にウォーレスから「変種がもとの型から無限に遠ざかる傾向について」という論文を受け取るまでは、図4-1のような樹形図を自身のノートに記すことはあったが、それを公表することはなかった。ところがウォーレスは、1855年に発表した最初の論文ですでに「共通の祖先型を設計図として2種ないしそれ以上の種が別々に形成されたときには、類縁性の系列は複合的なものとなり、フォーク状ないし多数に枝分かれした線でしか表現できない」と述べている。
◎共通祖先は化石として残っているか
それでは、共通祖先がどのようなものだったかを、われわれは知ることができるのだろうか。ラマルクには、共通祖先からの進化という明確な考えはなかったが、化石種は現生種の祖先だと考えていた。ところがダーウィンは、違っていた。
2つの近縁種を比較した場合、われわれはその共通祖先は両者の中間的なものと考えがちであるが、その通りであることはまずないという。少し極端な例かもしれないが、図4-3にある3種類の軟骨魚類(サメの仲間)を思い浮かべてみよう。

ほかのサメがすべて絶滅していたらという仮定は、極端だと思われるかもしれないが、似たようなことは実際に見られる。図4-4は、オーストラリアに生息するカモノハシとハリモグラを示している。これらの動物は単孔類と呼ばれる。

単孔類は、哺乳類でありながら、卵を産む動物である。単孔類には、昔はもっと多様なものがいたが、オーストラリアではカンガルーなどの有袋類が繁栄したために、単孔類の多くが絶滅したのだ。ハリモグラがなぜ生き延びられたかはよく分からないが、有袋類は水生適応するものがいなく(南アメリカには例外的に水生適応したミズオポッサムがいるが)、水中に進出したカモノハシは生き延びることができたのであろう。現在まで生き延びた単孔類のこの2つの系統は、形態的に非常に特殊化しており、彼らの共通祖先がどのようなかたちをしていたかを想像することは難しい。
たいていの場合、それぞれの系統での進化は異なる方向に進むので、個々の特徴について共通祖先が中間的であったということは、あまりないのだ。従って、化石種が複数の現生種の共通祖先であると判定することは、難しいのである。
さらに化石の記録は不完全だということがある。図4-5は、1938年に南アフリカのイーストロンドン沖のインド洋ではじめて捕獲された現生シーラカンスの写真である。

シーラカンスは化石としては古生代デボン紀から知られており、中生代白亜紀まで多くの化石が見つかっている。ところが、6,600万年前以降の新生代に入ると化石がまったく見つからないために、シーラカンスは恐竜と一緒に絶滅したと考えられていた。だから、シーラカンスがまだ生きていたという発見は大ニュースだった。シーラカンスがまだ生きているということは、恐竜絶滅以降も現在まで彼らの祖先が生き延びてきたということであり、化石証拠がいかに見つかりにくいかを示している。
ダーウィンが研究していたフジツボの仲間である蔓脚類では、彼の時代には新生代の化石で現生の属が知られているものは一つもなかった。彼が蔓脚類の論文を発表した直後に、新生代初期の地層からやっと一つだけ発見されたのだ。このことは、化石として発見されるのは、過去に生きていた生物のごく一部に過ぎないことを示している。従って、たまたま発見された化石が、現生種の共通祖先である可能性は極めて低いと言わざるを得ない。ダーウィンはキュヴィエと同様、化石種の大部分は絶滅した系統であろうと考えた。
キュヴィエは、動物界を図3-1のように4つの亜界に分類したが、亜界同士を結びつける中間型が見られないことから、ラマルクの進化論に反対した。その間を移行することは不可能だというのだ。
ダーウィンは、共通祖先から現生の亜界が進化するまでに、莫大な変化の積み重ねがあったために、現生の動物の比較だけではつながりが見えないのだと考えた。長い歴史を通じたつながりはあったが、化石記録の多くが現在に至る子孫を残すことなく絶滅したものであれば、そのつながりを見出すことは難しいのだ。
ダーウィンと同時代にイギリスで活躍した解剖学者にリチャード・オーエンがいた。彼はダーウィンの進化論に反対したことで有名であるが、比較形態学で多くの業績を残した。
ダーウィンがビーグル号航海から持ち帰った膨大な数の動物標本のうち、鳥類はジョン・グールドが調べたが、哺乳類、特に化石種については主にこのオーエンが調べたのである。オーエンが確立した概念のなかで、今日でも重要なものが「相同 Homology」である。これに対応するのが「相似 Analogy 」である。
今日では、「共通の祖先に由来する形質を相同」、「見かけ上は似ているが起源が違うもの(収斂)を相似」という。オーエンは進化を受け入れなかったが、ヒトの手と鳥の翼は相同であるが、鳥の翼と昆虫の翅は相似として区別した。このことは、直線的な発展ではなく、分岐の概念を受け入れる際の素地を与えるものと解釈できる。相同な構造は、基本パターンの変更に過ぎず、ヒトを目指して直線的に発展するという考えには反するものだからである。
キュヴィエやオーエンは進化論に反対したが、進化論の発展にとっては、彼らのような比較形態学のプロが抵抗勢力として頑張っていたということは重要であった。ダーウィンは『種の起源』を改定するたびに、自説への批判に対する自分の考えを書き加えることによって、進化理論の基盤を固めていったのだ。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


