
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第28話
ウイルスの突然変異率
文と写真 長谷川政美
◎突然変異率とゲノムサイズ
突然変異率を1回の複製で1塩基あたりに生ずる突然変異の割合とすると、一般にはゲノムサイズ(ゲノムの塩基数)の小さな微生物の突然変異率は大きくなる傾向がある。表28-1と図28-1がそのことを表している。
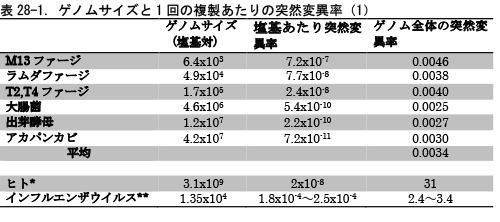
** 文献3
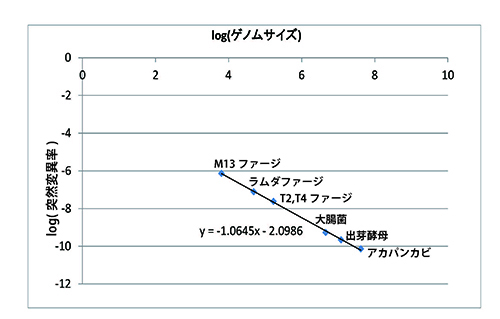
図28-1ではゲノムの大きな順にアカパンカビ(真菌)、出芽酵母(真菌)、大腸菌(真正細菌)、T2・T4ファージ、ラムダファージ、M13ファージが並んでいる。ファージは細菌に感染するウイルスのことで、バクテリオファージともいう。ファージとはギリシャ語で「食べる」という意味なので、バクテリオファージは「細菌を食べるもの」ということになる。
確かにゲノムサイズの大きな真菌から細菌、ウイルスとゲノムサイズが小さくなるにつれて1回の複製で1塩基あたりに生ずる突然変異は小さくなっている。この図は縦軸と横軸を対数で表しているが、きれいな直線関係が成り立っている。
このようにゲノムサイズと1塩基あたりの突然変異率との間で、きれいな逆相関関係が成り立つために、ゲノムサイズと1塩基あたりの突然変異率を掛け合わせたもの、つまり1回の複製あたりゲノム全体で生じる突然変異の数は、平均で0.0034とどの生物もほぼ一定の値になるのである(表28-1)。
なぜこのような逆相関関係が成り立つのであろうか。
たいていの突然変異は有害だから、もしも1回の複製当たりあまりたくさんの突然変異が生じるようだと、増殖のシステムが崩壊するという突然変異率の限界値があるのかもしれない。そうであれば、ゲノムサイズが大きくなるにつれて複製の精度が高くならなければいけないことになり、このような逆相関関係も成り立ち得るであろう。
しかし、そのようなことが文字通り成り立っているようには思えない。
その理由は、第一にゲノム全体の突然変異率が0.0034と、非常に低い値であり、これが限界値だとは考えにくいということにある(4)。第二に、ヒトなどの動物では明らかにこれとは違っているということである。また第三に、ウイルスだけに限っても、表28-1で取り上げたもの以外にも拡げてみると、逆相関関係が必ずしもきれいに成り立っているとは限らないことも分かってきた。
図28-1では、真核生物としては真菌類だけであるが、ヒトを入れると少し様子が違ってくる。ヒトの突然変異率は1塩基あたり1年あたり0.5x10-9とされている。1世代20年とすると、突然変異率は1塩基あたり1世代あたり2x10-8となる(表28-1)。これにヒトのゲノムサイズ31x108を掛け合わせると、ゲノムあたりの突然変異率は1世代で31と、微生物から得られた先の値0.0034のおよそ1万倍にもなる。
ヒトの場合、母親と父親から受け継いだ二揃いのゲノムをもつ(二倍体)ので、毎世代62個の新しい変異をもつことになる。ヒトやおそらくほかの動物も、図28-1の関係には従っていないことは明らかである。
その理由としては、ヒトのゲノムの大半は機能をもたない「ジャンクDNA」であるから、そこでたくさんの突然変異が起こっても構わないのだということが考えられる。1世代あたり1つのゲノムに平均31個の突然変異を生ずるということであるが、この推定値が正しいとすると、このうちの大部分が中立的な変異だと考えないと、生存の限界値を超えてしまうであろう。
しかし、近年ではいわゆるジャンクDNAの多くにも機能があるのではないかと考えられるようになってきた(5)。従ってヒトでゲノムあたりの突然変異率がなぜこのように高いのかについては、依然としてよく分からない問題である。
一方、ウイルスのようにゲノムサイズの小さなものでは、塩基あたりの突然変異率の増大がゲノムサイズの増大に制約を課している、あるいはゲノムサイズの増大が塩基あたりの突然変異率の増大に制約を課していると見られることがある。それが顕著なのが、ゲノムとしてDNAではなくRNAを使うRNAウイルスである。
◎RNAウイルスの突然変異率
図28-1に出てきたウイルスはすべてDNAウイルスであったが、一般にRNAウイルスはそれよりも高い突然変異率をもつ。細菌を含めて普通の生物は、ゲノムを複製する際に、複製エラーを修復する機構をもっている。ところがたいていのRNAウイルスには、この修復機構がないために、ほかのどのような生物よりも高い突然変異率をもつのである。
典型的な例がインフルエンザウイルスである。インフルエンザウイルスはオルトミクソ科に分類されるRNAウイルスであり、ゲノムはマイナス鎖の一本鎖RNAである。マイナス鎖とは、その情報が発現するためには一度転写されてプラス鎖の伝令RNAになる必要があるということである。一方、コロナウイルスのようなプラス鎖の一本鎖RNAウイルスでは、ゲノムがそのまま伝令RNAとして機能する。
宿主の細胞に感染したインフルエンザウイルスのマイナス鎖RNAはウイルス自身のRNA複製酵素とともに細胞核に入り込み、そこでプラス鎖の伝令RNAが転写され、さらにゲノムであるマイナス鎖RNAも複製される。その際に、多くの生物がもつ複製エラーの修復機構がないために、インフルエンザウイルスの突然変異率は非常に高いのである(6)。A型インフルエンザウイルスでは、1回の複製あたり1.8x10-4~2.5x10-4である(表28-1)。この値はDNAウイルスにくらべると非常に高い。インフルエンザウイルスのゲノムサイズは1.35x104とM13ファージとラムダファージの中間の大きさであり、ゲノム全体では1回の複製あたりの突然変異率は2.4~3.4になる。
インフルエンザウイルスを含めたたいていのRNAウイルスがゲノムを複製する際には、エラーが生じてもそれを修復する機構がないので、突然変異率が非常に高い。そのため、一人の患者のウイルス集団のなかでさえ、違ったゲノム配列が混ざっていることが多く、そのようなウイルスの集団を擬種quasi-speciesと呼ぶことがある(7)。
一本鎖RNAウイルスが変異を起こしやすいということは、ウイルスに対するワクチンを開発してもその効果を無効にしてしまうような変異が起こり得るということである。インフルエンザウイルスではそのようなことが起こるために、毎年新しいワクチン開発が必要である。
インフルエンザウイルスのような一本鎖RNAウイルスの多くは複製の際に起こるエラーを修復する機構をもっていないので突然変異率が高いが、同じ一本鎖RNAウイルスでもコロナウイルスは修復機構をもっている。そのためにコロナウイルスの1塩基座位あたりの変異率はインフルエンザウイルスにくらべると低いが、コロナウイルスは一本鎖RNAウイルスの中で最大のゲノムサイズをもっている。従ってゲノム全体で蓄積していく変異数でみると、かなりな数になり、感染の経路を追うのに使うことができるのである。
◎突然変異率とゲノムサイズ
SARS-CoV-2をはじめとして、近年問題になっているヒトの新興感染症ウイルスはすべてRNAウイルスである。これらのウイルスはもともと野生動物を自然宿主としていたものが、何かのきっかけでヒトにも感染できるように変異したものである。そのようなウイルスがすべてRNAウイルスだということは、RNAウイルスの突然変異率が高いことと関係していることは確かであろう。突然変異はランダムに起るが、そのような試行錯誤の回数が多ければ、たまたま出会ったヒトという新たな宿主に適合した変異が起こる確率は高くなるのだ。
逆に言うと、DNAウイルスは突然変異率が低すぎて新しい宿主に適応しにくい。従ってこれまでにうまく適応できた宿主のもとに留まろうとする傾向がある。SARS-CoV-2の祖先ウイルスのように、もともとキクガシラコウモリを自然宿主としていたものが短期間でヒトに感染できるように進化した上で、さらにヒトからヒトへと感染する能力をもつようになるなどということは、DNAウイルスには不可能なのだ。
ヘルペスウイルスは2本鎖DNAウイルスであるが、その中にヒトの水痘帯状疱疹(すいとうたいじょうほうしん)ウイルス(Human herpesvirus 3)がある。ヒトに適応したこのウイルスは、ヒトに感染すると最初は水痘(いわゆる「みずぼうそう」)を発症し、宿主が歳をとった数十年後に帯状疱疹として再び現れる。この間、ウイルスはヒトの細胞内に潜んで、ほとんど何もせずにじっとして宿主の免疫機能が低下して、再び活動できるようになるチャンスを待っているのだ。このように、DNAウイルスには一度感染できるようになった宿主にこだわった生き方を続ける傾向がある。
ただし、DNAウイルスが宿主を換えることがないということではない。ダニによって媒介される第26話で紹介したアフリカ豚熱ウイルスというDNAウイルスは、イボイノシシやアカカワイノシシから家畜のブタに宿主を換えた。これらの動物は互いに近縁なのである。
また、水痘帯状疱疹ウイルスに近縁のヘルペスBウイルスが自然宿主のアカゲザルにかまれたりしてヒトに感染することもあり、抗ウイルス薬ができるまでは致死率が非常に高かった。しかし、このような事故は実験用のサルを扱う施設で起るが、野生の環境ではたとえ密接な接触があった場合でも、サルからヒトにヘルペスBウイルスが感染して発症した例は知られていないという。
インドネシアのバリ島で観光客の与える餌目当てに集まるカニクイザルを調べたところ、成体の100%がヘルペスBウイルスの抗体をもっていたが、そこで働く観光業者でサルにかまれたり、引っかかれたりしたことのあるひとたちの中で、発症したひとはいないし、無症候で感染したひともいないのだ(8)。実験施設などでヘルペスBウイルスが稀な事故でサルからヒトに感染することはあるものの、自然界でサルからヒトに感染することはほとんどなさそうである。
一般にRNAウイルスのゲノムは小さいが、それはコピーを作るたびに生ずる突然変異率が高すぎて、ゲノムが大きいとエラーの総数が多くなって機能しなくなるためだ。ゲノムが小さいということは、複製のスピードが速く、短時間で大量のウイルス粒子が生産できるということである。
突然変異率が高いことが、ウイルス粒子数の多いことと相まって変異体を次々に生み出して新たな宿主の開拓を容易にしている。そのために野生動物からヒトに感染するようになって次のパンデミックを起こす候補として警戒の対象になるのは、RNAウイルスなのである。
【引用文献】
1.Drake, J.W., Charlesworth, B., Charlesworth, D., Crow, J.F. (1998) Rates of spontaneous mutation. Genetics 148, 1667-1686.
2. Scally, A., Durbin, R. (2012) Revising the human mutation rate: implications for understanding human evolution. Nature Rev. Genet. 13, 745-753.
3. Pauly, M.D., Procario, M.C., Lauring, A.S. (2017) A novel twelve class fluctuation test reveals higher than expected mutation rates for influenza A viruses. eLife 6, e26437.
4. Maynard Smith, J., Szathmáry, E. (1995) “The Major Transitions in Evolution”. W.H. Freeman.
5. 長谷川政美 (2020) 『進化38億年の偶然と必然』国書刊行会.
6. te Velthuis, A.J.W., Fodor, E. (2016) Influenza virus RNA polymerase: insights into the mechanisms of viral RNA synthesis. Nature Rev. Microbiol. 14, 479-493.
7. Moya, A., Holmes, E.C., González-Candelas, F. (2004) The population genetics and evolutionary epidemiology of RNA viruses. Nature Rev. Microbiol. 2, 279-288.
8. デビッド・クアメン(2021)『スピルオーバー:ウイルスはなぜ動物からヒトに飛び移るのか』甘糟智子訳、明石書店.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




