
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第14話
なぜコウモリを宿主とする
ウイルスが多いのか
文と写真 長谷川政美
近年、動物由来のウイルス感染症の相次ぐ出現が問題になっているが、それらのもとになった動物は「ウイルスの貯蔵庫」(reservoir of viruses)とも呼ばれる。前回、コウモリがウイルスの貯蔵庫として重要であることを論じたが、今回はこの問題をさらに考えてみよう。◎コウモリは特別な動物か
ヒトの感染症ウイルスのうちでコウモリに由来することがはじめて知られるようになったのが、1920年代に中南米のコウモリで見つかった狂犬病ウイルスであるが、その後たくさんのコウモリ由来のウイルス感染症が知られるようになったのは、この連載でこれまでに見てきた通りである。
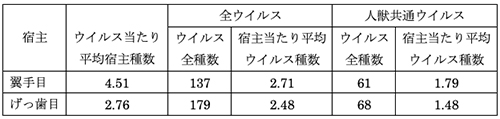
哺乳類5000種あまりのうちのおよそ20%が翼手目、42%がげっ歯目である。げっ歯目の種数は翼手目の2倍以上なので、目全体で知られているウイルスの数は翼手目よりもげっ歯目の方が多い。しかし、種あたりのウイルスの数では翼手目のほうが多いのだ(表14-1)(1)。しかもウイルスの中でヒトにも感染する人獣共通ウイルスに限ると、げっ歯目よりも翼手目一種当たりのウイルス種数がさらに多くなる傾向が見られる。
コウモリが多様なウイルスの貯蔵庫になっている理由の一つが、大きな集団で過密状態にいることが挙げられる(2)。メキシコオヒキコウモリ (Tadarida brasiliensis)は、一つのねぐらに100万匹が集まり、1平方メートル当たり3000匹の密度になる。これと近縁な図8-5で紹介したヒダクチオヒキコウモリも一つの洞窟に300万匹も密集して生息するという。
またコウモリの場合、一つの洞窟をねぐらとするのが一種だけではなく、いろいろな種類のコウモリが一緒に利用することが多い。そのために異種間での感染も起こりやすく、ウイルス当たりの平均宿主の種数がげっ歯目にくらべて翼手目が圧倒的に多くなっているものと思われる(表14-1)。

コウモリを自然宿主としているウイルスの中でヒトに感染するようになったものは、たいていは一本鎖RNAウイルスである。DNAウイルスにもコウモリを宿主としているものがあるが、その中でヒトに感染するようになったものは知られていない(3)。
新しい宿主に感染できるようになるためには、まず宿主細胞のもつ受容体とうまく結合できるようになることが必要であるが、一般に一本鎖RNAウイルスの突然変異率が高いので、種の壁を超えた感染を可能にするような変異が容易に生じやすいと考えられる(4)。
またたいていの場合、ウイルスはコウモリに対して病原性を示さない。これまでに知られているコウモリに感染するRNAウイルスの中でコウモリに対して強い病原性を示すものとしては、ジャマイカフルーツコウモリ(Artibeus jamaicensis)に致死的な症状をもたらすタカリベウイルス(Tacaribe virus)などわずかなものだけである(5)。このウイルスは、ラッサ熱を引き起こすラッサウイルスと同じアレナウイルス科(Arenaviridae)に属する。ジャマイカフルーツコウモリはこのウイルスの自然宿主ではなく、ほかの宿主から感染したものと考えられる。
ウイルスがコウモリに対して目立った病原性を示さないことに関連して、コウモリのゲノム解析から、彼らの免疫系の遺伝子は独自の適応的な進化を遂げたことが分かるという(6,7)。
図14-1に翼手目の系統樹を示す。翼手目は従来から小型の小コウモリと大型の大コウモリに分類されてきた。
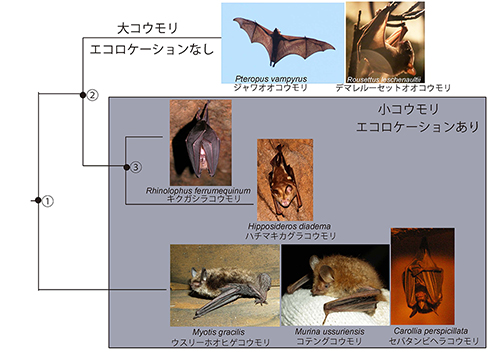
この分類はからだの大きさだけではなく、エコロケーションの能力の有無をも反映したものである。エコロケーションは反響定位ともいうが、動物が発する超音波の反響を受け止め、それによって周囲の状況を知ることである。
つまりコウモリの場合、飛びながら超音波を発してそれが昆虫などの獲物に反響するのを捉えて、夜間の暗闇でも獲物やまわりの状況をあたかも「見る」ように確認できる。レーダーである。夜行性の飛翔動物としてコウモリが成功したカギがエコロケーションにあるのだ。ただしエコロケーションの能力をもつのは小コウモリだけで、主に昼行性の大コウモリにはこの能力はない。
長い間、小コウモリと大コウモリという2大分類は、系統関係を反映するものと考えられてきたが、図14-1が示すように、分子系統学により同じ小コウモリでもキクガシラコウモリやカグラコウモリは、ホオヒゲコウモリ、テングコウモリ、ヘラコウモリなどよりもオオコウモリに近縁だということが明らかになってきた。
だとすると、エコロケーションは翼手目の共通祖先①ですでに進化しており、②で分岐したオオコウモリの系統で失われたという可能性と、①から分かれたあとのホオヒゲコウモリなどの小コウモリの系統と②から分かれた後の③に至る系統で独立に2回進化した可能性とが考えられる。オオコウモリにエコロケーションの痕跡があるという前者を支持する証拠と、小コウモリの2つの系統で聴覚に関与するたんぱく質で収れん進化が起ったという後者を支持する証拠とがあり、この問題はまだ決着がついていない。
超音波は高周波の声であり、コウモリはほかの哺乳類の発声と同じように喉頭から発する。その際、コウモリの口からは飛沫やエアロゾルが飛び散る。高周波音は急速に減衰するので、コウモリは大声を発し続けるのだ。アブラコウモリ属(Pipistrellus)が出す音は、1メートル以内の距離では120デシベルにもなることがあるという(9)。120デシベルとは、ジェット機のエンジン近くの音量であり、聴覚機能に異常をきたすといわれている。幸いにも、この音はわれわれの耳には聴こえないので、聴覚異常になる心配はない。コウモリに捕食される昆虫にもたいていこの音は聴こえないと思われるが、蛾の中にはこの音を聴きとって捕食から逃れる能力を進化させたものもいる。
このように小コウモリは飛沫をまき散らしながら飛び回るので、ウイルスも容易にほかの個体に感染し得るのである。しかもコウモリの寿命が長いことが、ウイルスの集団内での存続を助ける。さらにコウモリの飛翔能力はウイルスを遠くまで広めることにも貢献する。
このようなコウモリが抱えるウイルスがヒトに感染するようになったときに、重篤な感染症になる。そのためにコウモリの駆除がたびたび行なわれてきたが、それがさらなる混乱を引き起こしてきたことも第8話で見た。そこで述べたことと同じようなことがブラジルのチスイコウモリに対しても行われた(10)。チスイコウモリは家畜の血を吸うが、そのときに狂犬病ウイルスを感染させる。このために駆除の対象になったわけだが、ブラジル政府の援助のもとで行なわれたこの対策は、成果を挙げられなかっただけでなく、害虫を捕食したり、果物を食べて種子散布したり、送粉者の役割を果たすなどのさまざまな種類のコウモリも巻き添えにすることになり、生態系のバランスを完全に崩してしまったのだ。コウモリも統合された生態系の一員なので、それが欠けると生態系のバランスが保てなくなってしまう。
COVID-19の感染拡大もあり、多くの人々にとってウイルスは恐ろしいものだという認識があるかもしれないが、そもそもウイルスの中で病原性をもつものは限られている。ウイルスもまた統合された生態系の一員であり、その中でさまざまな役割を果たしていると考えられる。それぞれのウイルスが生態系の中で果たしている役割については、まだほとんど分かっていない状況であるが、彼らを欠くこともまた、生態系に対して予測できない影響を与えるであろう。第9話でも議論したが、近年の新興ウイルス感染症の相次ぐ出現には、人類が際限のない経済成長を求めて、生態系を攪乱させ続けていることに最大の原因があることを認識しなければならない。
◎宿主の大量絶滅とウイルスの絶滅
中生代白亜紀の終わりの6600万年前に恐竜が絶滅した。現在も繁栄している鳥類は恐竜の子孫だから、正確には非鳥恐竜が絶滅したというべきであろう。絶滅の原因は、その頃巨大な隕石が地球に衝突し、その衝撃によって起きた熱風や津波によるものだけでなく、衝突によって舞い上がった粉塵が太陽光を遮断したため、長期間にわたって地球全体が真っ暗闇になったからだとされている。そのような状況で植物も枯れてしまい、食料の尽きた非鳥恐竜が絶滅したのだ。これを白亜紀末大絶滅という。
この影響は、哺乳類や鳥類にも及び、彼らの多くも絶滅したと考えられる。しかし、彼らの多くはからだの小さな動物であり、小さな隙間などにわずかに残った食料を食いつないで生き延びることができたものもいたのであろう。いずれにしても、この大絶滅の時代を生き延びた系統であっても、大きな打撃を受けたことは確かであろう。
絶滅した非鳥恐竜や絶滅した哺乳類・鳥類を宿主としていたウイルスは絶滅した。なんとか生き延びた哺乳類・鳥類を宿主としていたウイルスはどうだったのだろうか。ウイルスは継続して感染を続けていかなければ生き延びられない。なんとか生き延びた宿主にしても、個体数やその密度は小さく、ウイルスが感染を維持するのは困難だったと思われる。隕石衝突は、宿主に対してよりも、ウイルスに対してより深刻な影響を与えたものと考えられる(11)。
翼手目が哺乳類のほかのグループから分かれて独自の進化の道を歩み始めたのは、今から7000万年以上も前のことだったと推定される(12)。そうだとすると、コウモリの祖先は6600万年前の白亜紀末大絶滅の時代を生き延びたことになる。およそ5250万年前の地層からオニコニクテリス(Onychonycteris)というコウモリの化石が発見されている(13)。この化石は現在のコウモリとそっくりであるが、まだエコロケーションの能力はもっていなかったようである。これとほとんど同時代に生きていたイカロニクテリス (Icaronycteris)という別のコウモリはエコロケーションの能力を獲得していたという。
大絶滅の時代、コウモリの祖先がどのような生活をしていたかは分からない。飛翔能力はまだ獲得しておらず、食虫性の小さな哺乳類として地表の小さな割れ目などで難を逃れたものと思われる。大絶滅に続いて鳥類など空を飛ぶような競争相手がまだ少なかった時代に飛翔能力を進化させたが、その後鳥類との競合もありエコロケーションが進化し、小コウモリは夜間の生活に適応していったものと思われる。
コウモリと共生する現在の多様なウイルス相はこのような過程で形成されたのであろう。コウモリとウイルスの間には、このような長い進化の歴史を通じて、宿主に対する病原性をほとんど表わすことのない共生関係が確立しているように思われる。ウイルスは宿主なしでは生きられないので、このような関係はウイルスにとって利点があるが、逆に宿主にとっても何らかの利点はあるのだろうか。次回はこの問題を考えてみよう。
1. Luis, A.D. et al. (2013) A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc. R. Soc. B280, 20122753.
2. Luis, A.D. et al. (2015) Network analysis of host–virus communities in bats and rodents reveals determinants of cross-species transmission. Ecol. Lett. doi: 10.1111/ele.12491.
3. Wang, L.-F., Anderson, D.E. (2019) Viruses in bats and potential spillover to animals and humans. Curr. Opin. Virol. 34, 79-89.
4. Mandl, J.N. et al. (2018) Going to bat(s) for studies of disease tolerance. Front. Immunol. 9, 2112.
5. Malmlov, A. et al. (2017) Serological evidence of arenavirus circulation among fruit bats in Trinidad. PLoS ONE 12(9), e0185308.
6. Jebb, D. et al. (2020) Six reference-quality genomes reveal evolution of bat adaptations. Nature 583, 578-584.
7. Nikaido, M. et al. (2020) Comparative genomic analyses illuminate the distinct evolution of megabats within Chiroptera. DNA Res. http://dx.doi.org/10.1093/dnares/dsaa021.
8. Teeling, E.C., Dool, S., Springer, M.S. (2012) Phylogenies, fossils and functional genes: the evolution of echolocation in bats. In “Evolutionary History of Bats” (eds. Gunnell, G.F., Simmons, N.B.), pp. 1-22. Cambridge Univ. Press.
9. J.D. オルトリンガム(1998)『コウモリ―進化・生態・行動』(松村澄子監訳)、八坂書房.
10. Meyen, F. (2003) Haematophagous bats in Brazil, their role in rabies transmission, impact on public health, livestock industry and alternatives to an indiscriminate reduction of bat population. J. Vet. Med. B50, 469-472.
11. Wang, L.-F., Walker, P.J., Poon, L.L.M. (2011) Mass extinctions, biodiversity and mitochondrial function: are bats ‘special’ as reservoirs for emerging viruses? Curr. Opin. Virol. 1,. 649-657.
12. dos Reis, M. et al. (2012) Phylogenomic datasets provide both precision and accuracy in estimating the timescale of placental mammal phylogeny. Proc. Roy. Soc. London B. 279, 3491–3500.
13. Simmons, N.B. et al. (2009) Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation. Nature 451, 818-821.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




