
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第22話
古い起源をもつウイルス
文と写真 長谷川政美
前回、細菌なみの大きさの核細胞質性大型DNAウイルス(NCLDV)が非常に古い起源をもち、真核生物が誕生した頃にこのウイルスの共通祖先が生まれた可能性があるという話をした。もしかして、このウイルスが真核生物の誕生と関わっているかもしれないのである。◎RNAポリメラーゼの系統樹
前回の図21-3の解析には含められていない遺伝子にRNAポリメラーゼ遺伝子(RNAP)がある。ググリールミニらはRNAPの分子系統学的な解析から、面白い仮説を提唱している(1)。
RNAPとは、DNA二本鎖の片方の鎖を読み取ってそれと相補的なRNAを合成する酵素であり、正式には「DNA依存性RNAポリメラーゼ」という。
コロナウイルスのようなRNAウイルスがRNAゲノムを転写してRNAを合成する際に用いるRNA依存性RNAポリメラーゼはこれとは違う酵素である。
真正細菌、古細菌、NCLDVなどはそれぞれ一種類のRNAPしかもたないが、真核生物は、DNAを鋳型にしてたんぱく質のアミノ酸配列情報を担う伝令RNA(mRNA)を転写するRNAポリメラーゼII (RNAP-II)だけでなく、リボソームRNAの転写を行なうRNAポリメラーゼI (RNAP-I)と転移RNA(tRNA)の転写を行なうRNAポリメラーゼIII (RNAP-III)など合計三種類のRNAPをもつ。
真核生物だけがなぜ複数の種類のRNAPをもつようになったかはこれまで謎であった。
それぞれのRNAPはおよそ12個のサブユニットで構成されている。その中で真核生物においてRPB1、RPB2と呼ばれているサブユニットは、真正細菌ではβ、β’、また古細菌ではB、Aと呼ばれているが、これらはすべてNCLDVにもあり、配列の相同性も明らかである。
つまりこれらはすべて共通祖先から進化したことになる。ググリールミニらがこの二つのサブユニットのアミノ酸配列を用いて系統樹を描いたところ、図22-1が得られた。
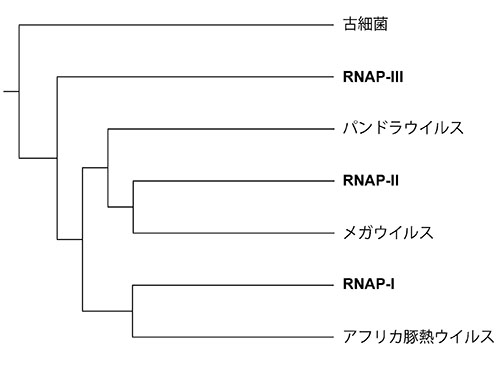
この系統樹が正しいとすると、なぜ真核生物が三種類のRNAPをもつようになったかについて、次のようなシナリオが描かれることになる(図22-2)。
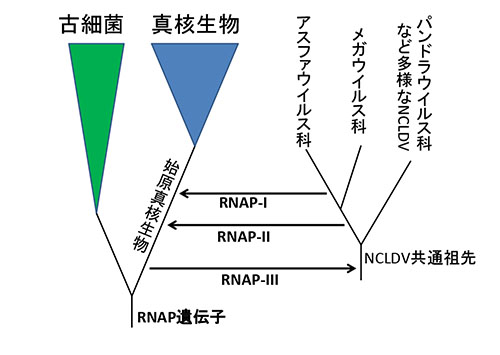
まず古細菌と真核生物の共通祖先は、一種類のRNAPをもっていた。真核生物ではあとでこれがRNAP-IIIに進化することになる。この遺伝子がNCLDVの共通祖先に水平伝搬した。水平伝搬というよりも、真核生物の祖先のゲノムから、RNAPとそのほかのいくつかの遺伝子をもとにして、新しいウイルスとしてNCLDVの祖先が誕生したのかもしれない。そうであれば、NCLDVはわれわれ真核生物の兄弟だということになる。
NCLDVは現在に至るまでこのRNAP遺伝子を保持し続けるが、進化の過程で少しずつ変異が蓄積した。アフリカ豚熱ウイルスを含むアスファウイルス科とメガウイルスの共通祖先から、始原真核生物と記された真核生物の共通祖先にこの遺伝子が水平伝搬し、真核生物ではその後RNAP-IIになった。
さらにアスファウイルス科がメガウイルスと枝分かれしたあとで、もう一度始原真核生物への水平伝搬が起こり、それがRNAP-Iになった。
真核生物が三種類のRNAPをもつのは、そのうちの二つをウイルスからもらったからだということになる。
このシナリオによると、NCLDVと始原真核生物の間の遺伝子のやり取りがあって、そのことが現在の真核生物、ひいてはわれわれヒトが進化するにあたって重要な役割を果たしたことになるのだ。
◎カウドウイルス
カウドウイルス目(Caudovirales)という真正細菌と共生するウイルスのグループがある。細菌のウイルスはファージと呼ばれるが、名前の「Caudo」がラテン語で「尾」を意味するように、尾部をもつファージである。二本鎖DNAの長さ18万~50万塩基対のゲノムをもつ。
あらゆる細胞性生物がもっている遺伝子の中で、ある種のウイルスももっていて、ウイルスの起源の問題と関連して注目されているのが先に出てきたDNA依存RNAポリメラーゼ(RNAP)である。
この酵素はたくさんのサブユニットからできているが、その中で先に出てきた真核生物のRPB1 とRPB2サブユニットに対応するものが真正細菌ではβとβ’サブユニットである。カウドウイルスもβとβ’をもつ。
実はRPB1 とβ、RPB2とβ’が相同であるだけでなく、対応しないサブユニット同士、つまりRPB1 (β)とRPB2(β’)の間でも相同性が見られるのである。このことは、この二つのサブユニットは、あらゆる生物の最後の共通祖先LUCAよりも前の段階で遺伝子重複によって生まれたことを意味する。
酸素を肺から全身に運ぶ役割を果たしているヒトのヘモグロビンというたんぱく質は、ヘモグロビンα鎖とβ鎖、それぞれ2本ずつ、合計4本のポリペプチドで構成されている。
このたんぱく質は、進化の過程をさかのぼるともともとは現在のミオグロビンのような1本鎖のたんぱく質であった。それがヤツメウナギと分かれたあとの顎をもった魚の段階で、遺伝子重複でα鎖とβ鎖が生まれて現在のヘモグロビンになったのである。このような遺伝子重複は、生物進化の至るところで起っており、新奇の機能が進化することに貢献してきたと考えられる(2)。RNAPの進化も、同じような遺伝子重複で起ったのである。
アメリカ・バージニア工科大学のアレイナ・ワインハイマーとフランク・エイルワードは、RNAPのこのサブユニットに着目した(3)。二つのサブユニットのアミノ酸配列を一つながりの配列として系統樹解析を行なったところ、図22-3のような系統樹が得られた。
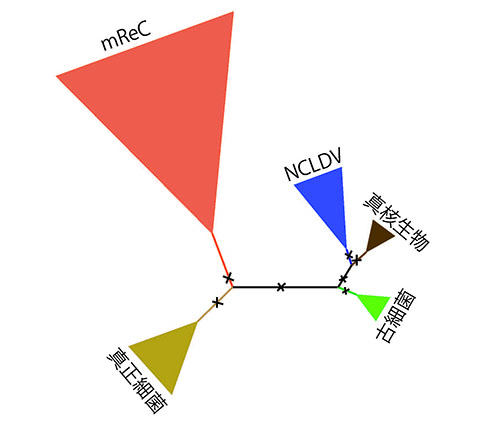
図22-3のような系統樹は無根系統樹と呼ばれるが、進化速度一定を仮定しない限り、一般の系統樹推定法では系統樹の根元の位置を決めることができない。生物学的には根の位置が重要な意味をもつが、それを決めるためには前提が必要である。
通常は、あるグループの生物種同士の系統関係を決めるためには、あらかじめそれらよりも遠い関係にあることが分かっているものを外群とし、当該グループ(内群)と外群を合わせた系統樹を推定したうえで、外群と内群の間に根があるとするのである。ところがこの図のような解析では外群にあたるものがないのだ。
このような問題を解決するための方法は、1989年にすでに得られていた。当時九州大学にいた岩部直之と宮田隆らが中心に行われ私も参加した研究で用いられた方法である(4)。
われわれは、真正細菌、古細菌、真核生物の間の系統関係を決める問題に取り組んでいた。ところが、この問題は地球上のあらゆる生物を含む系統樹を推定しようとするものだから、外群に相当する生物がいないのである。
進化速度が一定であれば外群なしでも系統樹の根の位置は決まるが、遺伝子の配列に相同性があって一つの系統樹にまとまる関係にあっても、これだけかけ離れた生物の間では進化速度が違うのが普通なので、外群なしで根の位置を決めることはできないのだ。
そこで岩部・宮田らが考えたのは、重複した二つの遺伝子を用いた「重複遺伝子の複合系統樹法」であった。あらゆる生物が共通にもっている重複遺伝子があれば、それらを合わせた複合系統樹を推定し、二つの重複遺伝子を結ぶ枝の途中に系統樹の根があるとすればよいだろうというわけである。
こうしてRNAPとは別の遺伝子だが、同様の重複遺伝子を用いて真正細菌よりも古細菌が真核生物に近縁だという結論が導かれた。長い間論争が続いていた生物系統学上の大問題に対して、解決の手掛かりが得られたのだ。
ワインハイマーらは、同じやり方でRNAPを構成するサブユニットの中の遺伝子重複で生まれた二つについて、複合系統樹解析を行なったのである(3)。彼らはカウドウイルス目のファージの中にもこれらのRNAPサブユニットをコードするゲノムをもつものがたくさんあることを見つけ、「mReC (multi-subunit RNAP-encoding Caudovirales」(複数のRNAPサブユニットをコードするカウドウイルス)と名づけた。カウドウイルスがすべてRNAPサブユニットをコードしているわけではないのだ。
彼らはほかの遺伝子を用いてまずmReCがカウドウイルスの中で系統的にまとまったグループを作ることを確かめた上で、複合系統樹解析にmReCカウドウイルスと核細胞質性大型DNAウイルスNCLDVも含めた。
結果は図22-4に示したが、NCLDVに関しては、先に紹介したように真核生物がさまざまな系統に分かれる前の始原真核生物から分岐した系統樹になっている。
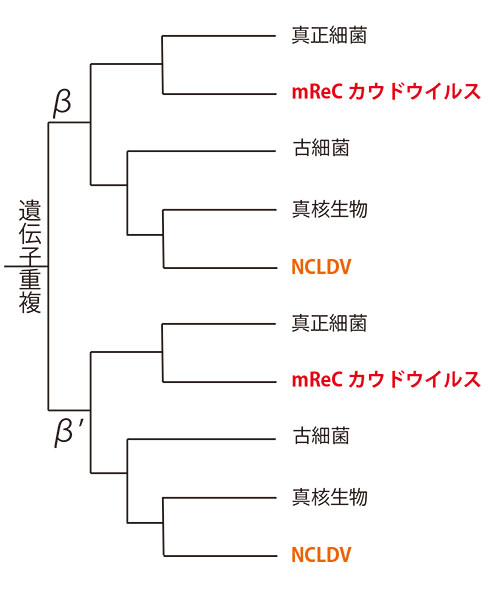
一方、mReCカウドウイルスはさまざまな系統に分かれる前の真正細菌の共通祖先から分岐している。つまりカウドウイルスもまた、非常に起源の古いウイルスの系統であり、真正細菌が現生のたくさんの系統に分かれる以前の共通祖先から、RNAPサブユニット遺伝子を水平伝搬で取り込んだ系統がmReCだということになる。というよりも、真正細菌の遺伝子が独立してカウドウイルスが生まれたのかもしれない。
図22-1の場合は、真正細菌を外群として系統樹の根元の位置を決めていると見なすことができたが、この場合は図22-2が示すように極端に進化速度が異なる系統が複数含まれるために、重複遺伝子の複合系統樹法の出番が必要な問題であった。
1. Guglielmini, J. et al. (2019) Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes. Proc. Natl. Acad. Sci. USA 116, | 19585-19592.
2. 大野乾(1977)『遺伝子重複による進化』山岸秀夫・梁永弘訳、岩波書店.
3. Weinheimer, A., Aylward, F.O. (2020) A distinct lineage of Caudovirales that encodes a deeply branching multi-subunit RNA polymerase. Nature Commun. 11, 4506.
4. Iwabe, N. et al. (1989) Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proc. Natl. Acad. Sci. USA 86, 9355-9359.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




