
いま世界に拡がるウイルスは、
リアルタイムで追跡できるほど進化が速い。ウイルスはいわゆる生物ではないが、
私たちヒトを含む生物と共進化する存在だ。
世界的な分子系統学者である著者が、
躍動感みなぎる“進化の舞台”へ読者を誘う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞を受賞。

ウイルスという存在
ーヒトの進化にどうかかわってきたのかー
第21話
生命の樹と巨大ウイルス
文と写真 長谷川政美
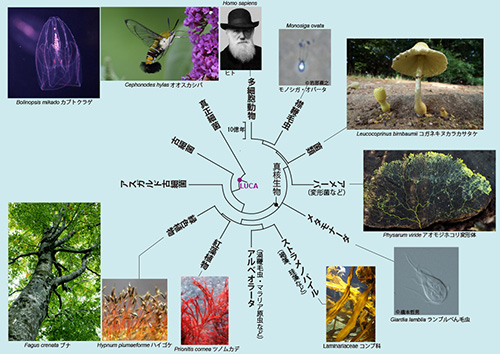
地球上のあらゆる生物は細胞からできており(そのため「細胞性生物」ともよばれる)、細胞の中にたんぱく質合成工場であるリボソームをもつ。リボソームはいくつかのリボソームRNA(主要なものは二つ)と多数のリボソームたんぱく質の複合体である。これらの大部分は、現存するあらゆる生物の最後の共通祖先であるLUCAがもっていたものである。リボソームは大小二つのサブユニットから成るが、小サブユニットの構成成分であるリボソームRNA(原核生物では16SrRNA、真核生物では18SrRNAと呼ばれる)は特に保存的であり、真正細菌、古細菌、真核生物の間で非常にはっきりとした相同性が認められる。
これ以外にも共通遺伝子はたくさんあるので、これらの遺伝子を用いて生物界全体をカバーする系統樹、「生命の樹」(Tree of Life)を描くことができる(図21-1)。これによりわれわれは、共通祖先LUCAから多様な生物が進化してきた様子を知ることができるのだ。
一方、ウイルスには細胞もなく、リボソームをもたない。彼らはゲノムをもつが、生物の細胞に入らないと活動できない。宿主細胞のリボソームを使って自身のゲノムにコードされたたんぱく質を合成してもらい、それを使って活動する。
細胞性生物のもつ遺伝子と共通の(相同性のある)遺伝子がウイルスのゲノムで見出されることもあるが、その由来がまったく不明の遺伝子も多い。しかも、あらゆるウイルスに共通した遺伝子がないので、生命の樹の中でウイルスがどのように位置づけられるか、不明なことが多い。また、はたしてあらゆるウイルスが一つの共通祖先から進化したものかどうかは、未だに答えが見つからない問題である。
すべてのウイルスに共通した遺伝子はないが、細胞性生物と共通の遺伝子をもつウイルスは多い。そのような遺伝子がウイルスの起源についての手掛かりを与えてくれるかもしれない。しかし、ウイルスと細胞性生物に共通した遺伝子は、ウイルスが細胞性生物のゲノムから取り込んだものかもしれないし(水平伝搬;図21-2)、逆に細胞性生物がウイルスから取り込んだものかもしれないので、必ずしもウイルスの起源に結びつくとは限らない。
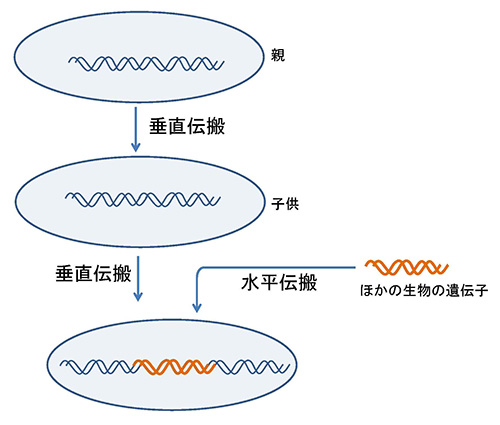
例えばこの連載のあとのほうで紹介するレトロウイルスなどは、生活環の中にプロウイルスとして自らのゲノムを宿主のゲノムに組み込むということがあるので、遺伝子を宿主のゲノムに残すことがある。そのようなことは、生物進化の歴史の中で比較的最近の出来事であり、ウイルスの起源と結びつくものではない。
◎巨大ウイルスの発見
イギリスのブラッドフォードという町で、1992年に流行性肺炎が蔓延した。その原因を突き止めるため、ある病院の冷却塔の水が検査されたが、その水の中からアカントアメーバ(Acanthamoeba)というアメーバが捕まえられた。アカントアメーバは図21-1に出てくる変形菌(粘菌ともいう)と同じアメーバ界に属する真核生物である。
実はこの時に光学顕微鏡でアメーバの細胞内に細菌のようなものが見つかり、「ブラッドフォード球菌」と命名されたのである。アカントアメーバも変形菌と同様に細菌を捕食するので、細菌が細胞内にいること自体は、珍しいことではなかったが、後にこれが間違いであることが判明した(1)。
実はこの「ブラッドフォード球菌」は、あらゆる細胞性生物がもっているリボソームRNAをもっていなかったのだ。ということは、これは細菌ではないことになる。
また、電子顕微鏡で調べると六角形のように見えた(立体的には20面体)。直径が0.75マイクロメートル(1000マイクロメートルが1ミリメートル)もあり、細菌の大きさに匹敵するものであるが、2003年になって結局ウイルスだということになった。それまでウイルスは光学顕微鏡では見えないと考えられていたので、最初はウイルスだとは思われなかったのである。
ウイルスの発見者としてマルティヌス・ベイエリンク(図1-1)と並んで取り上げられることの多いロシアのディミトリー・イワノフスキー(1864~1920)は、細菌を通さないような細孔の濾過器を通過できる「濾過性病原体」としてウイルスを発見したのである。細孔のサイズは0.2マイクロメートルほどなので、ウイルスはこれよりも小さいと考えられてきたのだ。
新たに発見されたブラッドフォードの巨大ウイルスは「ミミウイルス(Mimivirus)」と命名された。この名前はmimic(模倣する)からきているが、細菌に似ているということである。
ミミウイルスのゲノムは二本鎖DNAである。このウイルスは巨大だというだけでなく、ゲノムも大きく、120万塩基対もある。大腸菌のゲノムサイズは460万塩基対でこれよりも大きいが、同じ真正細菌でもマイコプラズマは58万塩基対と小さいし、もっとゲノムの小さな細菌もいる。ミミウイルスのゲノムがコードする遺伝子の数は、980個にもなる。
さらにミミウイルスがウイルスらしくないのは、この遺伝子の中にtRNA(転移RNA)遺伝子とともに、アミノアシルtRNA合成酵素の遺伝子が4個含まれていることである。アミノアシルtRNA合成酵素とはtRNAにアミノ酸を結合させる酵素であり、アミノ酸をつけたtRNAがリボソームにやってくることによってたんぱく質の合成が進む。
普通のウイルスはこのような遺伝子はもたずに、宿主のものを使う。ただし、アミノ酸は20種類あるので自分の遺伝子だけでまかなおうとすると、20種類のアミノアシルtRNA合成酵素の遺伝子が必要になるが、ミミウイルスはアルギニン、システイン、メチオニン、チロシン用の4種類しかもたない。
ミミウイルスはリボソームRNA遺伝子などをもたないので自前の遺伝子でリボソームを作ることはできないが、なぜアミノアシルtRNA合成酵素遺伝子の一部だけ中途半端なかたちでもっているのであろうか。
大きさとゲノムサイズがミミウイルスよりもさらに大きなメガウイルス(Megavirus)は、ミミウイルスがもつ4種類に加えてさらに3種類のアミノアシルtRNA合成酵素の遺伝子をもつ。これらのウイルスはもともと20種類のアミノアシルtRNA合成酵素の遺伝子をもっていたが、それを失いつつある進化(退化)の途上にあるように思われる。
その後見つかったパンドラウイルス(Pandoravirus salinus)のゲノムは、さらに大きく247万塩基対もあり、2556個ものたんぱく質をコードする遺伝子をもつ。しかし、これらの遺伝子のうちの2155個は、データベース中に相同な配列が見つからないという。つまり遺伝子の由来が分からないのだ。由来の分かった遺伝子のうちの2個がアミノアシルtRNA合成酵素のものであり、それはミミウイルスやメガウイルスなどほかの巨大ウイルスの遺伝子よりも宿主であるアカントアメーバの遺伝子に近いという。
先にミミウイルスやメガウイルスなどでアミノアシルtRNA合成酵素の遺伝子が少数ではあるが見つかるのは、これらのウイルスの共通祖先が20種類の遺伝子セットをもっていたものが、進化の過程で失いつつあるように見えると述べた。パンドラウイルスはこれとは逆に、宿主から新たに遺伝子を水平伝搬で取り込んでいるようなのである。
◎核細胞質性大型DNAウイルス
海洋のメタゲノム解析により巨大ウイルスが次々に見つかるようになった。メタゲノムとは、ウイルスを培養せずに環境中のゲノムを解析する方法である。ゲノム以外のウイルスの実体は捉えられなくても、このような解析によりさまざまなウイルスが海洋中にいることが分かってきた。その中には、ミミウイルス、メガウイルス、パンドラウイルスなどに似た二本鎖DNAゲノムをもつ巨大ウイルスたくさんいたのである。
水深5~50メートルの海の表層近くでは海水100 mlあたり平均450万個、水深300メートルでは7~23万個の巨大ウイルスの存在が明らかになった。巨大ウイルスは決して珍しいものではなく、海洋環境で普通に存在していたのである。また陸上でも節足動物、哺乳類、両生類、爬虫類など決して特殊とはいえない環境でも見られる。
これらのウイルスはいくつかの遺伝子を共有しており、系統的に一つのグループを形成している可能性が高まってきた。このグループは「核細胞質性大型DNAウイルス」(Nucleo-cytoplasmic large DNA virus; NCLDV)と呼ばれる。このグループは大型で真核生物にしか感染せず、細胞核内あるいは細胞質で複製を開始し、最終的に細胞質で複製を終えることから命名されたものである(2)。
◎巨大ウイルスの起源は古い
核細胞質性大型DNAウイルスNCLDVにはさまざまなものが含まれる。赤潮の原因となるラフィド藻(Heterosigma akashiwo)に感染する巨大ウイルスもその一つだが、赤潮を抑制する働きがあるという(2)。ラフィド藻は、図21-1の中の褐藻を含むストラメノパイルに属する不等毛植物である。
実は、有名な天然痘ウイルスもNCLDVとしては比較的小さな36万塩基対のゲノムサイズにもかかわらずNCLDVの仲間らしいが、まだ不明な点もある。このほかに、ダニを介してブタに感染するアフリカ豚熱ウイルスも同じ仲間である。
NCLDVは本当に系統的に一つのグループを作っているのだろうか。個々の遺伝子は水平伝搬によって細胞性生物とウイルスの間やウイルス同士で交換されることがあるので、個々の遺伝子の系統樹解析だけではこの問題を解明することはできない。
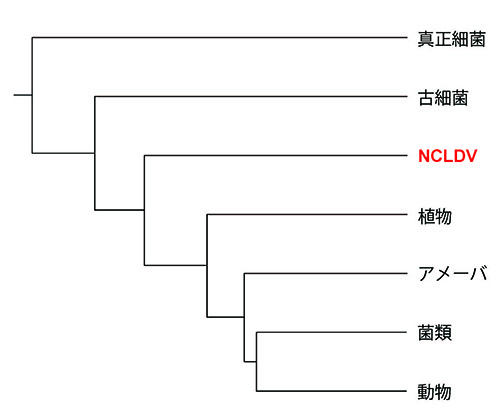
フランス・パスツール研究所のジュリアン・ググリールミニらのグループは、96種のNCLDVが共通にもっている保存的なたんぱく質遺伝子8個を選び出し、細胞性生物の相同遺伝子とあわせて系統樹解析を行なった。8個のたんぱく質の示す系統関係はほぼ同じものであり、NCLDVの進化的な関係を示すものと考えられる。それが図21-3の系統樹である(3)。それによると、NCLDVは系統的に一つにまとまっているとともに、真核生物が植物や動物などさまざまな系統に分かれる前の真核生物系統樹の根元から分かれている。
図21-1では、真核生物は古細菌の中のアスガルド古細菌から分かれているが(つまり古細菌は系統的にまとまったグループではない)、ググリールミニが所属するパスツール研究所のパトリック・フォルテ―ルらのグループはそれを認めない(4)ので、図21-3ではアスガルド古細菌を含めていない。アスガルド古細菌はほかの古細菌にはない真核生物と共通の特徴をもつが、系統樹解析に用いる遺伝子を取り換えると、アスガルド古細菌が真核生物に特に近縁とはならないという。しかしいずれにしても、真核生物誕生の頃に、NCLDVも誕生しているのだ。
1.武村政春(2015)『巨大ウイルスと第4のドメイン』講談社.
2.三原知子、五斗進、緒方博之(2015)海洋巨大ウイルス―ゲノムから見えてきた多様性と生態.生物の科学 遺伝 69 (4), 318-325.
3.Guglielmini, J. et al. (2019) Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes. Proc. Natl. Acad. Sci. USA 116, | 19585-19592.
4.Da Cunha, V. et al. (2018) Asgard archaea do not close the debate about the universal tree of life topology. PLoS Genet. 14, e1007215.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『ウイルスとは何か:生物か無生物か、進化から捉える本当の姿』 (中公新書)。ウイルスは恐ろしい病原体か、あらゆる生命の源か――。進化生物学の最前線から、その正体に迫る。

☆いちばん新しい本!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




