
ヒトとチンパンジーの共通祖先は600万年前に生きていた。
この地球上に、ヒトとゾウの共通祖先は9,000万年前、
ヒトとチョウの共通祖先は5億8,000万年前、
ヒトとキノコの共通祖先は12億年前に生きていた。
15億年前には、ヒトとシャクナゲの共通祖先が生きていたという…。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。復旦大学生命科学学院教授(中国上海)。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)『遺伝子が語る君たちの祖先―分子人類学の誕生』(あすなろ書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。

僕たちの祖先をめぐる15億年の旅
第14話
カエルとヒトの共通祖先
文と写真 長谷川政美
前回は、鳥類の祖先をたどり、ジュラ紀から白亜紀にかけて全盛を誇った恐竜についてお話ししました。さらにその祖先をたどると何に出会うでしょうか? 現在生きている動物のなかで鳥類に一番近いのは爬虫類のワニ類ですから、次に出会うのはワニとの共通祖先○19です(下図をクリックして確認してください)。
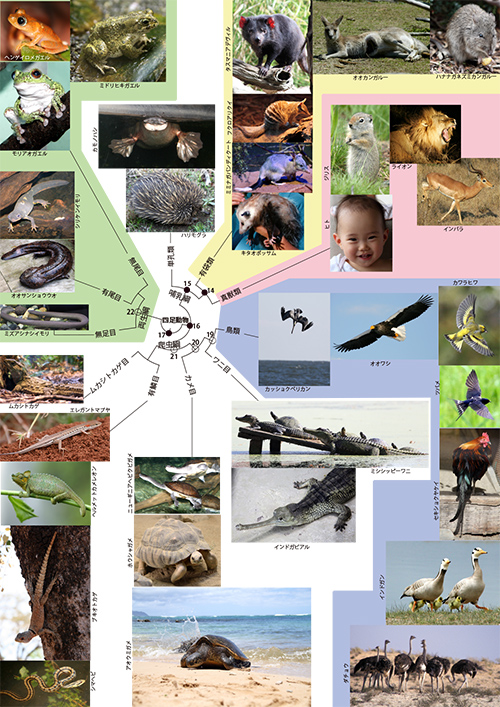
現生のワニには、上図(図11-1)の四足動物系統樹マンダラにあるミシシッピーワニやインドガビアルのように、4本の足をからだの外側に突き出して、地面を這うように歩くというイメージがあります。そもそも爬虫類の「爬」は、地面を這うという意味です。トカゲも似たような歩き方をします。
ところが、鳥が地面を歩くときは2足歩行で、足はからだの下の位置で体重を支えます。恐竜も2足歩行が多いですが、大型草食恐竜のように4足歩行のものでも、4本の足がからだの下に位置して体重を支える歩き方をします。その点で、現在のワニは恐竜よりもトカゲなどに似ているようにも思えます。

ところが、三畳紀のワニ(図14-1)は、恐竜と同様に足がからだの下で体重を支えていました。このように足をからだの重心の下にして体重を支えながら、まっすぐ伸びた足を前後に動かしながら移動するのは、トカゲのように地面を這うよりも効率がよいと考えられます。恐竜(もちろん鳥類も)や哺乳類はみなそのような歩き方をします。
現生のワニがトカゲに似た姿勢に見えるのは、水辺で餌をとる半水棲の生活に適応したためだと考えられます。さらにワニの祖先とされている三畳紀のテレストリスクスは、鳥類のように2足歩行をしていたようです。
また、現生のワニでも、休んでいるときは足をからだの外側に突き出していますが、歩くときには地面に垂直に足を伸ばしてからだを地面から持ち上げた状態になります。
多くのワニは、鳥のように、胃の筋肉部分である砂嚢にためた小石を使って飲み込んだ肉を腸に送るのに都合のよい大きさにまで砕いて小さくします。さらに、ワニは卵を守り子供の世話をするなど、現生の爬虫類のなかでは鳥類に近い習性をもっています。
鳥類+恐竜とワニ類をあわせて「主竜類」と呼びますが、○19(図11-1)の子孫がすべて主竜類です。ところで現生のワニはすべて肉食性ですが、中生代白亜紀にはワニ類はもっと多様で、草食性と考えられるものもいました(図14-2)。

鳥類+恐竜の姉妹群がワニ類であることは、以前から言われていましたが、鳥類+恐竜+ワニ類の姉妹群が何かについては現在とは違う考えが受け入れられていました。
爬虫綱にはワニ目以外にトカゲやヘビなどの「有鱗目」(これは真獣類のセンザンコウが属する目と同じ名前ですが、大きく離れたグループの場合は混同する恐れがないので両方とも使われています)、「ムカシトカゲ目」、それに「カメ目」があります。そのなかで有鱗目+ムカシトカゲ目が鳥類+恐竜+ワニ類の姉妹群であり、カメ目はさらに遠い関係にあるとされてきたのです。
この考えの最大の根拠が側頭窓の数でした。頬の壁には顎を閉じる筋肉である側頭筋がありますが、顎を閉じるときにこの筋肉は収縮し、そのために外側に膨れ出ようとします。筋肉が収縮すると顎のなか空間に入りきらなくなるのです。従ってこの筋肉が発達すると、収縮した筋肉をはみ出させるために、側頭窓という孔が頭骨の両側にあいたとされています(図14-3)。
哺乳類型爬虫類や哺乳類では、1つの孔が頬にあいているため、これらの動物を「単弓類」と呼びます。

一方、ムカシトカゲ目とワニ目それに鳥類、恐竜では2つの側頭窓があり、これらは「双弓類」といいます。ヘビ・トカゲなどの有鱗目では少し変形したこめかみをもっていますが、2つの側頭窓をもつ型に由来すると考えられるので、これらも双弓類です。ところが、カメ目は、魚類や両生類と同じように側頭窓をもたないため「無弓類」と呼ばれます。
このようにカメは祖先型の特徴をもっていることから、爬虫類のなかでは最初に他から分かれたものと考えられたのです。ところが、分子系統学は鳥類+ワニ目の姉妹群はカメ目であって、ムカシトカゲ目+ワニ目はもう1段階遠い関係にあることを示したのです。ですから、側頭窓の有無は系統関係を明らかにする際の手掛かりにはならないのです。
.jpg)
こうして鳥類の祖先をたどる旅は、ワニ目との共通祖先○19(図11-1)、カメ目との共通祖先○20と次第にさかのぼり、有鱗目+ムカシトカゲ目との共通祖先○21にたどり着きます。○21はすべての爬虫類の共通祖先ということです。恐竜が爬虫類であるならば、鳥類も爬虫類の一員ということで差支えないでしょう。
◎鳥類とヒトの共通祖先
ここで久しぶりに再び僕たちヒトの祖先をたどる旅に戻りましょう。哺乳類の共通祖先●15(図11-1)をさかのぼると、鳥類を含む爬虫類の共通祖先○21の祖先でもある●16に出会います。●16から進化したものはすべて「羊膜類」と呼ばれます。鳥類を含む爬虫類と哺乳類が羊膜類です。爬虫類や単孔類の卵は、胚が羊膜という膜でおおわれ、そのなかに満たされた羊水のおかげで、胚が乾いてしまわないようになっています。胎生哺乳類の真獣類や有袋類でも、胎児を包んでいる構造は同じようなものです。このような羊膜のおかげで、羊膜類の動物は、カエルなどの両生類のように卵を水中に産まなければならないという制約から解放され、一生を陸上で過ごすことができるようになったのです。
●16は、僕たち哺乳類と鳥類の共通祖先ですが、現在生きている哺乳類と鳥類のどちらにも似ていたとは考えられません。現在生きている爬虫類もすべてこの祖先から進化したのです。ですから●16は外温性で卵のかたちで子供を産んでいたと思われます。
◎海に戻った羊膜類
陸上生活に適応した羊膜類のなかで、海に戻ったものもたくさんいます。哺乳類のクジラ、ジュゴン、爬虫類のウミガメなど数え切れないほどです。そのなかで、ダーウィンが訪れたことで有名なガラパゴス諸島の海イグアナについてお話しましょう。
ガラパゴスは南アフリカの太平洋沖の島々で、300~400万年前に火山の噴火でできました。ここには海イグアナ以外に陸イグアナがいます。南アメリカから海を越えてやってきた陸イグアナのなかから海藻を求めて海に戻ったのが海イグアナです。ところが、DNA解析でこの2種のイグアナが分かれた年代を推定すると1,000~2,000万年前と島ができた時代よりも前になってしまいます。これは一時進化学者の間で大問題になりましたが、真相は以下の通りです。
現在のガラパゴス諸島は確かに新しいのですが、実は2,000万年前にはすでにその付近に火山島があったのです。最初の陸イグアナがその島にやって来て、そこで海イグアナが生まれました。その島はその後海中に沈みましたが、両方のイグアナはその前に新たにできた島に移住したのです。
◎最初に陸上に上がった僕たちの祖先
共通祖先●16(図11-1)をさらにさかのぼると両生類との共通祖先●17に出会います。もともと生命は海で生まれましたが、陸地は長いあいだ生命のない世界でした。なぜなら、初期の地球には紫外線が大量に降り注いでいたからです。紫外線は遺伝子DNAを壊すため、生物にとって有害でした。水は紫外線を吸収するので、生物にとって水中での生活は安全でした。
シアノバクテリアや植物が光合成で酸素を出すにつれて、次第に大気中の酸素濃度が上昇し、それにつれてオゾンが増えていきました。オゾンは酸素原子が酸素分子と結びついてできるものですが、有害な紫外線を吸収します。そのために、地上に降り注ぐ紫外線の量は減っていったのです。
およそ3億6,000万年前になると、陸上でも脊椎動物が住める環境が整えられてきました。最初に陸上に上がった脊椎動物が両生類です。ですから共通祖先●17は、僕たちの祖先が両生類のような生活をしていた頃に相当します。側頭窓もまだできていなかったことでしょう。●17から進化したものはすべて「四足動物」と呼ばれます。二本足の鳥類も僕たちヒトも四足動物の一員です。祖先が4本足で陸上を歩いていたからです。
僕たちとカエルの共通祖先●17が、3億6,000万年前に陸上に進出する以前に、地衣類、植物、昆虫などはすでに陸上に進出していました。食料としてのそれらを追って、僕たちの祖先は川や沼から陸上に移住したのです。
◎足が退化した四足動物
現在の両生類は3つの目(もく)に分類されています。カエルなどの「無尾目」(オタマジャクシには尾はありますが、おとなのカエルには尾がないので)、イモリ、サンショウウオなどの「有尾目」、アシナシイモリなど「無足目」です。
四足動物系統樹マンダラ(図11-1)にあるアシナシイモリは足がなくてまるでミミズのような外見ですが、背骨をもったれっきとした脊椎動物です。足が退化するという現象は、四足動物のさまざまな系統で独立に起りました。両生類のなかでもアシナシイモリ以外に有尾目のサイレンというグループ(図14-4)、爬虫類のなかでヘビ以外にも足のないトカゲが何回も進化しています(図14-5)。これも一種の収斂です。


ヘビは4本の足を完全に失いましたが、細長い体をくねらせることによって地上、樹上、水中などでの移動が可能であり、さまざまな環境に適応しました(図14-6)。
鳥類では前足が翼になったわけですが、空を飛ばなくなった鳥のなかには、キーウィーのようにさらに翼を完全に失ったものもいます(図14-7)。


*もっと詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本連載に大幅な加筆をして、新たな図版を掲載したものです。
扉絵:小田 隆
ブックデザイン:坂野 徹


