
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第47話
鳥類の台頭と翼竜の衰退
文と写真 長谷川政美
◎植物食恐竜の体重分布
前回、植物食恐竜で特に体重の分布が右に偏ることをみた。哺乳類でも生態的には植物食恐竜に近いと思われる植物食に特化した偶蹄目について体重の種数分布を調べてみると、図47-1のようになる。前回、図46-4に示した齧歯目と翼手目の分布と違って、図46-5の鳥盤類や竜脚類の分布ほど顕著ではないが、これらに少し近いものになっている。従って、右に偏った(左に裾を引いた)分布は、鳥盤類や竜脚類などの非鳥恐竜に限ったものではないのかもしれない。
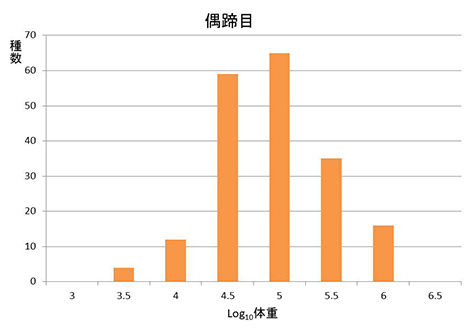
図47-1. 偶蹄目における体重の対数と種数頻度の関係。ただし、クジラ、カバ、ブタなどは食性がかなりほかの偶蹄目と違っているので除いた。データベースPanTHERIA http://esapubs.org/archive/ecol/E090/184/により作成。
非鳥恐竜の体重分布の偏りを考えるにあたって、もう1つ重要なことは彼らの産んだ卵がからだの割に小さいということである。第17話で取り上げたマダガスカルの絶滅した走鳥類エピオルニス・マキシマスの卵は、いかなる巨大な恐竜の卵よりも大きかったという。従って、成体は巨大な竜脚類であっても、卵から孵化した幼体は小さかったはずである。そのために、彼らは成長の過程でさまざまに異なったニッチを占めていたものと考えられる。つまり、大型恐竜の幼体が小型恐竜のニッチを占めていたために、もともと小型恐竜の進化が抑えられていた可能性がある。
オゴルマンとホーンはさらに、非鳥恐竜の体重分布を時代ごとに調べてみた(図47-2)。この場合、分類群ごとに分けるとサンプル数があまりにも少なくなるために、すべての非鳥恐竜はまとめて調べた。
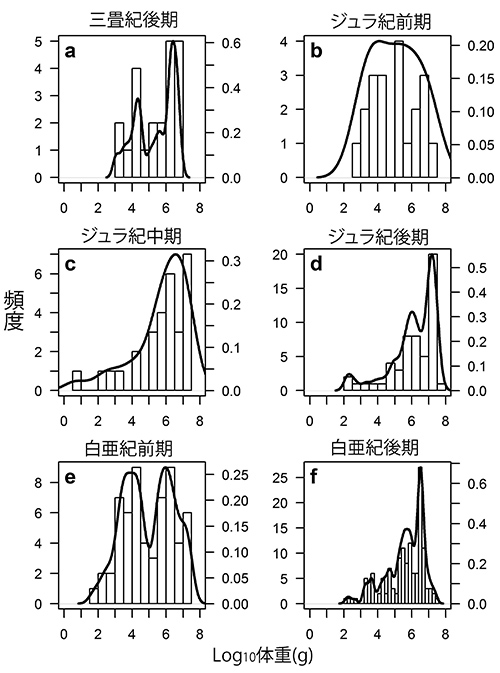
図47-2. 非鳥恐竜の時代ごとの体重分布の比較(O’Gorman and Hone, 2013)。
その結果、ジュラ紀前期と白亜紀前期では、あまり偏りのない体重の分布を示した。それに対してジュラ紀後期と白亜紀後期では、体重の平均値よりも最頻値が右に偏るような分布になっているのだ。
ペニーらが「白亜紀の末期に小型恐竜が衰退していた」と言っているのは、このことに対応するのかもしれない。もしそうであれば、白亜紀後期だけでなくジュラ紀後期にも似たような傾向があるにもかかわらず、非鳥恐竜がそのまま絶滅しないで白亜紀にも繫栄していたことに対する合理的な説明が必要である。
いずれにしても、このような問題に関わる化石データはそれほど豊富に得られるわけではないので、小型恐竜が白亜紀末期には衰退していたといっても、それが本当の傾向を示したものなのか、あるいはサンプリング誤差による見かけ上のものに過ぎないのかを見極めることが難しい。今後もっとたくさんの化石データを使った解析が可能になって、果たして白亜紀後期に小型恐竜の衰退が本当に起っていたのかどうかを検定できるようになることが望まれる。
たとえペニーらが言うように「小型恐竜が白亜紀末期には衰退していた」のが事実だとしても、白亜紀末期は大型恐竜進化の絶頂期だったように見える。白亜紀末期まで繁栄していたティラノサウルスは、肉食恐竜としては大きさの点では必ずしも最大ではなかったが、それでも体長12メートルを誇り、動物史上最も強い噛む力をもっていたと言われている。歯は大きなもので長さ30センチメートルもあり、その2/3は歯根として顎の骨に埋まっていたので、しっかりと獲物に噛みつくことができた。
さらに、北海道大学の小林快次さんらの研究によると、脳の構造から、ティラノサウルスの仲間は臭球という臭いを感知する脳の一部の大きさが、ほかの獣脚類恐竜よりも発達していたという。従って、臭いを感知して獲物を探し出す能力もすぐれていたのだ。
このように高度に進化した捕食者の脅威にさらされていたトリケラトプスやエドモントサウルスなどの大型植物食恐竜も、彼らが強力な捕食者と長い期間共存していたということは、それぞれに高度な進化を遂げていたはずである。
巨大隕石が、これら進化の絶頂期にいた非鳥恐竜らを滅ぼしたことは確かであろう。通常は絶頂期から衰退がはじまるというが、非鳥恐竜の場合は、絶頂期に突然絶滅したように見えるのだ。
◎翼竜の衰退
中生代には非鳥恐竜とともに翼竜が栄えていた。これまで脊椎動物のなかで空に進出したのは、鳥類、コウモリ、それに翼竜だけである。そのなかで、空中に進出することに最初に成功したのが翼竜であった。それは今からおよそ2億2000万年前の中生代三畳紀後期のことだった。
翼竜は皮膜を使って飛行する。皮膜を使って自力で飛行するように進化したのは、翼竜以外にはコウモリしかいないが、皮膜を滑空に使う動物はたくさんいる。ヒヨケザル、ムササビ、トビトカゲなどである。皮膜が少しでも発達すれば、樹上から飛び降りる際に、滞空時間を延ばし、目標場所に狙いをつけて着地するのに役立つであろう。従って、最初に皮膜をもった翼竜の祖先は、捕食者から逃れたり、獲物をとる上で有利だったであろう。
このように、翼竜の皮膜は最初から飛行のために使われたと考えられる。それに対して、鳥類が飛行に使う羽毛は、もともと飛行の役に立っていたとは考えにくい。羽毛は、広く恐竜の仲間で保温やディスプレイに使われていたものが(図47-3)、鳥類の祖先で2次的に飛行の用途に使われるようになったと考えられる(第27話)。

図47-3. 中国遼寧省で最初に見つかった羽毛恐竜:シノサウロプテリクスSinosauropteryx prima(白亜紀前期; 中国科学院南京地質古生物研究所所蔵)。首から背筋にそって繊維状の構造がみとめられ、これが羽毛の原型だという。
翼竜は、非鳥恐竜とともにK/Pg境界(6600万年前)の時期に絶滅するが、白亜紀末にはケツァルコアトルスに代表される巨大な翼竜しか残っていなかったという(図47-4)。2016年に、翼開長がおよそ1.5メートルの小型翼竜の化石が白亜紀末の地層から見つかり話題になっているが、K/Pg境界以前からすでに小型翼竜が衰退していたことは確かである。なぜ彼らが衰退したかというと、鳥類との競合が有力な原因と考えられる。

図47-4.白亜紀末期まで北アメリカにいたケツァルコアトルスQuetzalcoatlus northropiの骨格標本(北九州市立・いのちのたび博物館所蔵)。翼開長がおよそ10mと推定される史上最大の翼竜。日本語では恐竜や翼竜の名前はたいていカタカナで表記されるが、中国語ではケツァルコアトルスは「風神翼龙」と呼ばれる。
シギ、チドリなど現在水辺に生息する鳥を「渉禽類(しょうきんるい)」という。白亜紀にはまだシギ、チドリの仲間は出現していなかったが、彼らと同じような生活をしていた鳥はいた。水辺を歩き回る鳥や小型翼竜は足跡を化石として残しやすい。
ジュラ紀には翼竜の足跡化石がたくさん見つかるのに対して、白亜紀に入って鳥の足跡化石が現れると、翼竜のものは少なくなり、白亜紀最後のマーストリヒチアン期(7210~6600万年前)にはなかなか見つからなくなるのだ。
このことから、白亜紀に入ってからの鳥類の急速な進化が、小型翼竜に大きな影響を与えたと考えられる。ジュラ紀までの翼竜には翼開長が2.5メートルを超えるものはいなかったが、白亜紀前期になると翼開長が6メートルを超えるものが現われ、白亜紀後期になると大型のものだけになったのだ。
白亜紀を通じてまず小型翼竜が衰退し、白亜紀末期にはケツァルコアトルスのように巨大な種が少ししか残らなくなったということは、ペニーらが考えた非鳥恐竜の衰退パターンと似ている。巨大な種が進化するための源である小型翼竜の衰退によって、翼竜全体が衰退していったように思われる。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


