
知の巨人アリストテレス、分類学の父リンネ、
古生物学の創始者キュヴィエと連なる、自然に対する知識を体系化する博物学は、
19世紀半ばにダーウィンとウォーレスの進化論に到達した。
事実に基づき、歴代の学者たちが打ち立てた仮説の数々を丁寧に読み解きながら、
分子系統学の登場で新たな時代を迎えた“進化学の現在”までを追う。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)『新図説 動物の起源と進化―書きかえられた系統樹』(八坂書房)など多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。進化が一目でわかる「系統樹マンダラ」シリーズ・ポスターは全編監修を務める。

進化の歴史
ー時間と空間が織りなす生き物のタペストリー
第42話
分解者を食べる変形菌の進化
文と写真 長谷川政美
◎物質循環の抑制と生態系の多様化
2億9900万年前の石炭紀と古生代最後のペルム紀の境界付近で、植物遺体を完全に分解できるハラタケ綱の菌類が進化した。こうして、ぺルム紀のあいだに生態系の物質循環がうまく回るようになってきた。
2億5100万年前にぺルム紀が終わり、中生代の三畳紀に入ると陸地の生態系は古生代にくらべてはるかに豊かになってきた。しかし、ここでこの豊かな生態系を支えるもう1つの立役者を紹介しておかなければならない。それが変形菌(粘菌ともいう)である。
生態系の物質循環がうまく回るためには、生産者(植物)、消費者(動物)、分解者(菌類、細菌)の存在が重要であるが、多様な生物が生きていくためには物質循環の速度も重要である。
落ち葉や倒木などは、小さな生き物に住み場所を与えている。これらが菌類や細菌によってすみやかに分解されてしまったら、それを住み家にしている小さな生き物たちは生活の場を失うことになる。さらに地面の保水効果も弱まって、乾燥化が進んでしまうだろう。
物質循環がうまく回るためには、分解者の存在は必須であるが、あまり速く回っても多様な生物が生きていくことはできない。変形菌が菌類や細菌といった分解者を食べることによって、結果として落ち葉や倒木の分解速度が抑えられているのだ。
◎変幻自在な変形菌
変形菌には菌という名前がついているが、菌類(真菌類)とは別物である。菌類は系統的には動物(多細胞動物)に近縁であり、この2つをあわせてオピストコンタという。変形菌は、真核生物の系統樹・図8-2で出てきたアメーボゾアに属するものであり、アメーボゾアはオピストコンタとは別の系統であるが、アメーバは動物のように動き回ることができる。
今回出てくる変形菌はすべて、「~ホコリ」という名前をもつが、風が吹いたり動物が触ったりすると、乾燥した子実体から名前にある「ホコリ」のように胞子が大量に飛ぶからである。昔は「~ホコリカビ」と呼ばれていたが、菌類ではないために、現在では「~ホコリ」という。
変形菌は一般のひとにはあまり馴染みがないかもしれないが、少し注意すればわれわれの身のまわりで普通に見られる生き物である。図42-1a、b、cにどのような場所で変形菌が見られるかを示した。
特に日本では梅雨の時期によく見かける。その時期に、図42-1 a、b、cのような倒木の上や木の切り株などを注意するとよい。ただし、変形菌は一般には小さいものなので、その美しさを堪能するには、ルーペなど拡大して見るものが必要になる。肉眼では見過ごしてしまうが、ルーペをのぞくと、思いがけなく美しい小さな世界が出現する。これらの写真で見えている変形菌は、すべて子実体という胞子を作る時期のものである。
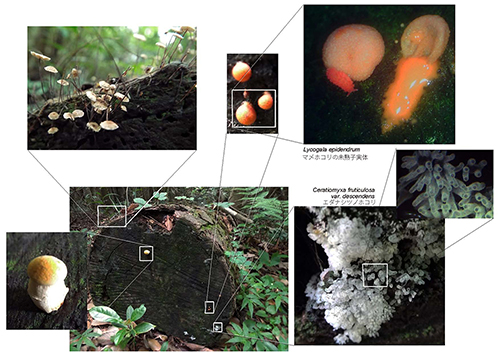
図42-1a.変形菌はこの写真のように、倒木や木の切り株、落ち葉の上などで見ることができる。日本では梅雨の時期によく見かける。この写真(6月10日)では、マメホコリとツノホコリの子実体が見える。右上のマメホコリの子実体は、未熟で水分を多く含んでおり、つぶすとこのようにネバネバの液体が出る(右の子実体)。近くには左側にあるような様々な菌類も見られる。クリックすると大きく表示されます。

図42-1b.朽ちた倒木の上に生えたPhysarum sp.モジホコリとCeratiomyxa fruticulosa var. descendensエダナシツノホコリ(6月29日)。クリックすると大きく表示されます。

図42-1c.切り株に生えたTubifera ferruginosaクダホコリの未熟な子実体(6月29日)。
変形菌の一生は胞子の発芽から始まる(図42-2)。胞子が発芽すると、およそ10マイクロメートル(0.01ミリメートル)ほどの単細胞のアメーバ(粘菌アメーバ)が現れる。
このアメーバは最初自分よりも小さな細菌を食べて大きくなる。アメーバにはオス・メスがあり、異性が出会うと接合し、染色体数が2nの変形体になる。変形体でも、核は分裂して増えるが細胞は分裂せずに単細胞のまま大きくなる。次第に大きな変形体になると、大きな菌類も食べて成長する。時には1メートルの大きさになることもある。変形体は動物のように動き回り、餌となる菌類や細菌を探す。
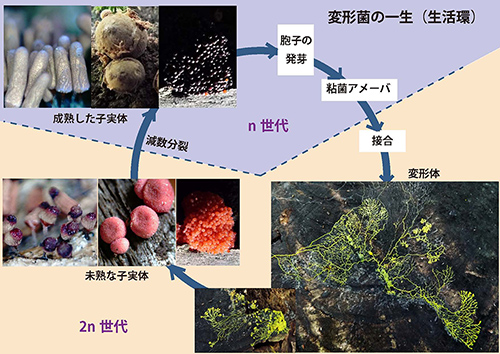
図42-2.変形菌の一生(生活環).変形体と子実体の写真は、必ずしも同じ種のものではない.子実体の一番右の写真はクダホコリTubifera ferruginosaであるが、真っ赤な未熟子実体が翌日行ってみると黒くなっていた(一部赤い部分も起っているが).未熟な子実体が成熟する過程で減数分裂が起り、染色体数nの胞子が作られる。クリックすると大きく表示されます。
図42-3に木の切り株上に出てきた変形体を示した。このような広がったもの全体が、1つの細胞なのだ。左上の変形体を撮影したおよそ3時間後が左下の写真であるが、そのあいだに変形体のかたちが変わり、移動していることがわかる。翌朝見ると、変形体はほとんど消えて、代わりに子実体が現われていた。
変形体の移動は、細胞の中身の原形質流動によって引き起こされる。この流動を起す原動力は、動物の筋肉と同じようなアクチンとミオシンの相互作用によっている。アメーバが細菌や菌類を食べる際には、第33話で紹介した食作用により、食べ物を食胞で包み込んで細胞内に取り込み、その中で消化する。
一方、菌類などはキチンなどで作られた細胞壁をもつために、食作用はできないので、木材や地中に広く張り巡らされた菌糸から消化酵素を体外に分泌して分解された養分を吸収するのである。このように、変形菌と菌類は大きな食べ物から栄養を摂るために、それぞれ違った仕組みを進化させた。

図42-3.木の切り株上に出てきた変形体。それぞれ部分を拡大した写真も示した。左上の写真のおよそ3時間後に撮影したのが左下の写真.変形体のかたちが変わり、移動したのが分かる。右が翌朝の様子。変形体はわずかに残っているだけだったが、切り株の表面をよく見ると、アオモジホコリPhysarum virideと思われる多数の子実体が生えていた。この子実体は未熟で湿っているのでみずみずしい緑色であるが、成熟するにつれて次第に黄色に変わる。さらに次の朝の状態が、右の一番上の写真である。胞子を飛ばす準備の整った成熟した子実体になっていた。クリックすると大きく表示されます。
図42-4にこのほかのさまざまな変形体を示した。変形体はその後、胞子をつくる子実体になる。子実体ができても、図42-5のように、周りに変形体が這いまわった跡が残っていることがある。
変形菌は粘菌とも呼ばれるが、動物のように動き回るアメーバや変形体の時期は水分を含んでねばねばしているからである。餌となる菌類や細菌は湿気の多い場所に多いので、変形体は倒木の中や落ち葉の裏側で成長する。
次世代に命を繋ぐための胞子をつくる子実体は乾燥した場所で形成される。そのため変形体は明るい場所に這い出てくる。そこで作られる子実体は最初カラフルなものが多い(図42-2)。このような未熟な子実体は乾燥するにつれて成熟した子実体になる。成熟する過程で減数分裂が起り、染色体数がnの胞子が作られる。

図42-4.さまざまな変形菌の変形体。南方熊楠は変形体を「混沌たる痰のような」と形容した。変形体は動き回って菌類や細菌を食べる。これはたくさんの核をもった一つの巨大な細胞であり、単細胞生物の究極の生き方といえる。菌類・担子菌(図41-2)の菌糸は枝分かれしながら広がるが、互いに癒合することはなくて網状にはならないのに対して、変形体の広がった脈同士は癒合して網状になる。(d)は子実体が形成されつつあるところ。

図42-5.Fuligo septica var. flavaキフシススホコリの子実体の周りに変形体が這いまわった跡が見られる。
図42-6は、子実体の写真を用いた変形菌の系統樹マンダラである。広い意味の変形菌は、真正粘菌亜綱のほかに、ムラサキホコリ亜綱、ツノホコリ亜綱を含む。この図でも水分を含んだカラフルな未熟子実体が乾燥するにつれて色が変わる様子がわかる。風に乗って胞子を飛ばすためには、まず胞子を乾燥させることが必要なのである。このように子実体は乾燥の過程で色が変わるが、変形菌はかたちが変わるだけでなく、色も変化するのだ。

図42-6.変形菌の系統樹マンダラ.子実体の写真を用いて描いた.同じ種の写真で(a)、(b)とあるものは、その順で未熟で水分の多いものと乾燥が進んだものである。クリックすると大きく表示されます。
日本で見られる変形菌は、およそ500種と言われている。一方、世界中の変形菌はおよそ1,000種である。世界中の変形菌のおよそ半分もの種が日本で見られるのだ。このようなことは他の生物では考えられない。これには、日本の湿気の多い気候が関係していると考えられるが、変形菌の胞子は小さく、世界中に散布されやすいために世界共通の種が多いことの反映でもある。


図42-7.小動物によって食べられる変形菌.(a)変形菌の変形体を食べるダンゴムシ、(b) Tubifera ferruginosaクダホコリの未熟な子実体を食べるヤスデ.このほか、甲虫、トビムシ、カタツムリなども特に変形菌の子実体を好んで食べる.彼らの移動距離は短いが、成熟した子実体を食べることによって胞子の拡散に貢献しているかもしれない。
◎変形菌研究の先達たち
変形菌の変形体は時には1メートルにもなる大きな細胞であるが、変形体を2つに切ると、流れ出た原形質が固まって、遺伝的に同等な2つのアメーバ個体になる。多細胞動物のプラナリアも2つに切ると、それぞれが再生して2個体になるが、変形体がプラナリアと違うのは、分かれた2つが出会うと再び一つの個体に戻ることである。動物のように動き回っていた変形体が、キノコのように胞子を作る子実体に変身するが、子実体が成熟する過程でさまざまな色彩の変化が見られる。
昭和天皇と南方熊楠は、このような変幻自在な変形菌(粘菌)の魅力に惹かれた研究者であった。昭和4年(1929)に熊楠の住む和歌山県田辺への昭和天皇の行幸があった。6月1日熊楠は、田辺沖の神島(かしま)で陛下に拝謁した。唐澤太輔さんによると、小雨の降る中およそ25分間、神島で陛下は変形菌を採集されたという。その後、田辺湾に停泊中の御召艦・長門で熊楠の御進講が行われた。25分間の予定だったが、陛下の希望で5分か10分延長されたという。そのとき、熊楠は変形菌などの標本をキャラメルの空箱に入れて献上したというエピソードが残っている。翌年には、神島に熊楠がこの島の森が末永く保たれることを願って詠んだ和歌の石碑が建立された(図42-8a)。昭和16年(1941)に熊楠は亡くなったが、その後、昭和37年(1962)、再度の行幸の折りに、昭和天皇は熊楠を偲ぶ和歌を詠まれた。この御製を刻んだ碑は、神島を望む、田辺市の南方熊楠記念館の前に建立されている(図42-8b)。

図42-8.南方熊楠(1867-1941)と昭和天皇(1901-1989)は、ともに変形菌の研究者として知られている。昭和4年(1929)に熊楠の住む和歌山県田辺への昭和天皇の行幸があり、熊楠は田辺沖の神島で陛下に拝謁した。(a)は翌年に神島に建立された熊楠の和歌の石碑、(b)は昭和37年(1962)に再度の行幸の折りに、昭和天皇が熊楠を偲んで詠まれた和歌の石碑である。
*もっと「進化」を詳しく知りたい人に最適の本:
長谷川政美著『系統樹をさかのぼって見えてくる進化の歴史』 (ベレ出版)。 本書は当サイトで連載していた「僕たちの祖先をめぐる15億年の旅」を加筆修正および系統樹図を全て作り直して一冊にまとめたものです。カラー図版600点掲載。
扉絵:小田 隆
ブックデザイン:坂野 徹


