
870万種ともいわれる地球上の多様な生き物たち。
まだ私たちはそのごく一部しか知らないが、
実に多くのことが明らかにされてきてもいる。
進化生物学者である著者が、
世界中で長年撮りためた貴重な写真と文章で
思いのままに「生き物」を語る。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

進化の目で見る生き物たち
第12話
動物のからだの模様の形成
文と写真 長谷川政美
多くの動物のからだにはさまざまな模様が見られる(1)。前回はネコ科の進化を紹介したが、図12-1にそのなかのヒョウ属の系統樹とそれぞれの動物の模様を示した。
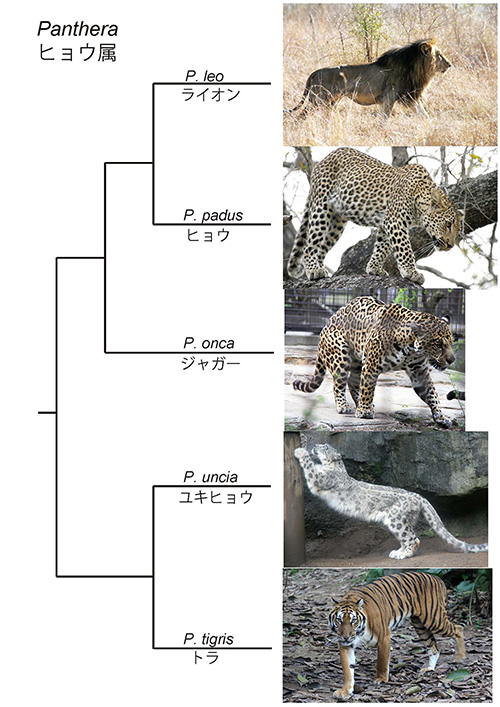
ノーベル文学賞を受賞したイギリスの作家ラドヤード・キプリング(1865~1936年)が自分の子供を寝かしつけるために語っていた物語をまとめた童話集『なぜなぜ話』(2)のなかで出てくる、シマウマやヒョウの模様は彼らの生息環境では自分の存在を目立たなくしているという話が代表的な答えである。しかし、このような話は実証することが難しく、さまざまな議論がある。今回は、このような問題は避けて、どのような仕組みで模様が作られるかという点について考えてみよう。
◎チューリングのモデル
多細胞動物のからだは、一個の受精卵から出発して、細胞分裂を繰り返しながらできてくる。もともと一個の細胞が分裂するだけであれば、ある細胞は表皮に、別の細胞は消化器系に、さらに別の細胞は神経系にと分化するのはなぜか、という疑問がわく。
どの細胞も同じゲノムをもっているのに、細胞はそれの属する組織によって異なる形質をもつのはなぜなのだろうか。それは、ゲノム中のすべての遺伝子が働いているのではなく、必要な遺伝子が必要な時期だけ働くように制御されていており、この制御が組織によって異なるためである。
発生過程において、このように遺伝子に働いて発現を促す物質をモルフォゲン(morphogen、形態形成物質ともいうが、かたちだけでなく色素の発現などに関わる物質も含む)という。細胞の分化は、モルフォゲンが細胞ごとに不均一に分布することによって起こると考えられる。しかし、もともと同じ細胞なのに、モルフォゲンが不均一に分布するのは、どのようにしたら可能なのだろうか。この問題は、からだの表面にできる模様のできかたにも関わる。動物のからだの模様は、色素を作る細胞が不均一に分布することによって作られるからである。
現在のコンピュータ科学の基礎をつくったイギリスの天才的数学者アラン・チューリング(1912~1954年)は、亡くなる2年前の1952年にこれを説明するモデルを提唱した(4)。
私は50年ほど前にこのモデルを紹介する記事を書いたことがあるが(5)、ここではそれを極力簡略化して再度紹介しよう。高校の数学程度の知識で理解していただけると思うが、数式アレルギーのかたは数式の部分は読み飛ばしていただいても、文章と図だけである程度理解していただけるだろう。
チューリングは、XとYというモルフォゲンを考え、それらの濃度(それぞれxとyで表わす)が、一つの細胞内で、次のような微分方程式に従って時間的(t: 時間)に変化するとした。
![]() (1a)
(1a)
![]() (1b)
(1b)
このような系は、次のような場合に実現される。
1) X、Yとも一定速度1で
![]() (2)
(2)
![]() (3)
(3)
2) Yは7yの速度で壊れていく。
3) Xは6xの速度でYに変換される。
4) Xは自分自身の生成反応を
![]() (4)
(4)
5) XがYの濃度に依存して6yの速度で減っていくような機構がある。
つまり、モルフォゲンXは自分自身の合成を促進し、さらにモルフォゲンYの合成も促進する。またYはXの合成を止める。この系の定常状態はx = y = 1のときである(このとき、式(1)でdx/dt、dy/dtがゼロになる)。
図12-2に示すように、定常状態(1, 1)から攪乱してずらしてやっても、またもとに戻るから、この状態は安定である。ところが、このような閉じた反応系ではなく、反応系が2つあって、その間でX、Yが拡散によって行き来できるとすると、様子がまったく違ってくる。
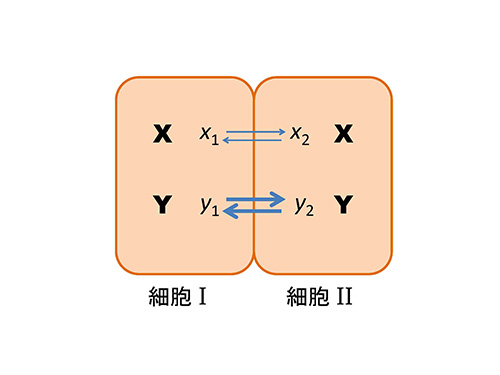
図12-3のように2つの接触し合った等価の細胞Iと細胞IIを考える。

![]() (5a)
(5a)
![]() (5b)
(5b)
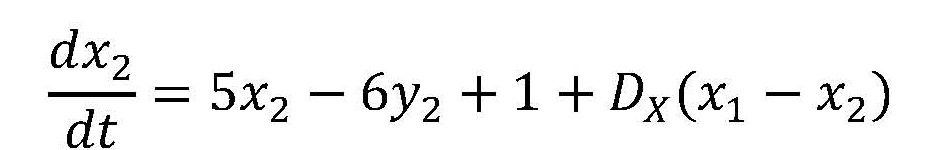 (5c)
(5c)
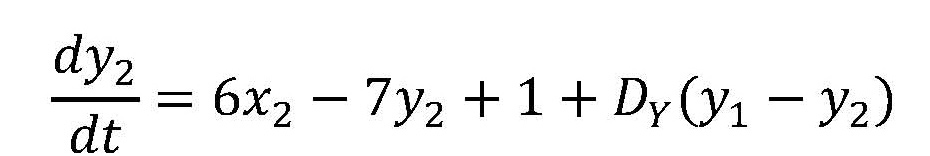 (5d)
(5d)
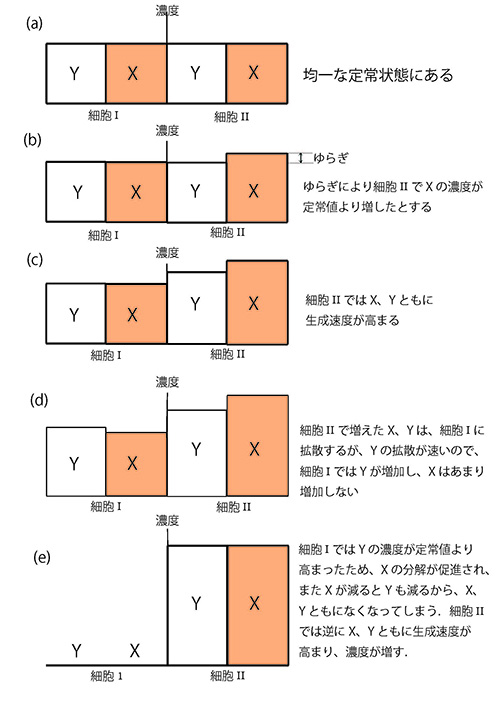
このように細胞IではYが定常値よりも高まった結果、Xの分解が促進され、さらにXが減るとYも減るから、結局X、Yともになくなってしまう。一方、細胞IIのほうでは、逆にX、Yともに生成速度が高まり、濃度が増す(e)。こうして、2つの細胞の間で誘導物質X、Yの不均一な分布が実現される。
このようにして、モルフォゲンが2つの細胞の間で不均一に分布する結果、この2つの細胞は別々の方向に運命づけられ、分化が起る。ここでは、2つの細胞系について議論したが、このことは必ずしも2細胞期の発生段階でこのようなことが起るという意味ではない。発生のいろいろな段階でこのような仕方で細胞の分化が可能だということである。
チューリングのこの反応・拡散系のモデルではYがXにくらべて十分速く拡散するということが重要である。そのようなことは、Xが高分子、Yが低分子の場合に実現する。
◎繰り返しをもった模様の形成
これまでチューリングのモデルを使って、2細胞系で分化が起ることを説明したが、もっとたくさんの細胞の系では、モルフォゲンの濃度が定常波のようになる。これを「チューリング波」という。チューリング波ができることによって、繰り返しのある模様、つまりヒョウの斑点模様やシマウマの縞模様が作られている可能性があるのだ(6,7)。
最近の研究では、このような繰り返しのある模様のできる仕組みが遺伝子レベルで解明されつつある(8)。その結果、チューリングが70年前に考えたモデルと文字通り同じものではないが、似たような反応と拡散を組み合わせた系が、斑点や縞のような繰り返しをもった動物の模様の形成に寄与していることが明らかになってきた。
チーターは図12-5aのように斑点模様をもつが、南アフリカの限られた地域ではキングチーターと呼ばれる変異体が見られる。キングチーターでは図12-5bのように斑点が大きく、さらにそれが連なっている(特に背中では完全に連なっている)。このような模様の違いも遺伝子レベルで解明されつつある(9)。


◎甲虫の模様
図12-6にオオキノコムシ科の2種の甲虫の写真を示した。この2つは似た4つのノコギリ状の紋様をもち、同じ科だが別属に分類される。しかし、同じ属のものがみな同じような模様をもつとは限らない。さらに、このようなノコギリ状の紋様をもつものは、ゴミムシダマシ科(10)、シデムシ科(図12-7a)、オサムシ科(図12-7b)、テントウムシダマシ科(11)など鞘翅目のさまざまな科で見られる。このように同じような紋様がさまざまな系統で見られることは、この紋様は容易に進化しうるものであって、紋様形成には何か共通の機構が働いているものと推測される。






【引用文献】
1. 寺井洋平、米澤隆弘、長谷川政美(2012)斑な世界を形作る遺伝子たち.ビオストーリー 18, 28-37.
2. ラドヤード・キプリング(1902)『ゾウの鼻が長いわけ ― キプリングのなぜなぜ話』(日本語:藤松玲子訳、2014年、岩波書店).
3. Li, G., Davis, B.W., Eizirik, E., Murphy, W.J. (2016) Phylogenomic evidence for ancient hybridization in the genomes of living cats (Felidae). Genome Res. 26(1), 1-11.
4. Turing, A.M. (1952) The chemical basis of morphogenesis. Phil. Trans. Roy. Soc. B237, 37-72.
5. 長谷川政美(1972)閉じた系と開いた系.現代化学 1972(11), 16-26.
6. 近藤滋(2019)『波紋と螺旋とフィボナッチ』角川書店.
7. 近藤滋(2021)『いきもののカタチ ― 多彩なデザインを創り出すシンプルな法則』学研プラス.
8. Kaelin, C.B., McGowan, K.A., Barsh, G.S. (2021) Developmental genetics of color pattern establishment in cats. Nature Comm. 12, 5127.
9. Kaelin, C.B., Xu, X., Hong, L.Z., et al. (2012) Specifying and sustaining pigmentation patterns in domestic and wild cats. Science 337, 1536-1541.
10. 秋田勝己、安藤清志、平野雅親、柏原精一、益本仁雄、大澤省三、吉川寛(2013)「科の壁」を超えて - 摩訶不思議なゴミムシダマシの多様性.月刊むし506, 13-27.
11. 大澤省三(2015)多様性に満ちた甲虫の進化.『遺伝子から解き明かす昆虫の不思議な世界』(大場裕一・大澤省三・昆虫DNA研究会編、悠書館)、pp. 347-408.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。




