
870万種ともいわれる地球上の多様な生き物たち。
まだ私たちはそのごく一部しか知らないが、
実に多くのことが明らかにされてきてもいる。
進化生物学者である著者が、
世界中で長年撮りためた貴重な写真と文章で
思いのままに「生き物」を語る。

著者プロフィール
長谷川政美(はせがわ まさみ)
1944年生まれ。進化生物学者。統計数理研究所名誉教授。総合研究大学院大学名誉教授。理学博士(東京大学)。著書に『分子系統学』(岸野洋久氏との共著)『DNAに刻まれたヒトの歴史』(共に岩波書店)、『系統樹をさかのぼって見えてくる進化の歴史』(ベレ出版)、『世界でいちばん美しい進化の教室』(監修、三才ブックス)、『共生微生物からみた新しい進化学』(海鳴社)、『進化38億年の偶然と必然』(国書刊行会)など多数。進化に関する論文多数。1993年に日本科学読物賞、1999年に日本遺伝学会木原賞、2005年に日本進化学会賞・木村資生記念学術賞など受賞歴多数。全編監修を務める「系統樹マンダラ」シリーズ・ポスターの制作チームが2020年度日本進化学会・教育啓発賞、2021年度日本動物学会・動物学教育賞を受賞。

進化の目で見る生き物たち
第4話
海を越えた動物の移住
文と写真 長谷川政美
前回は、海流に乗って運ばれた果実がほかの大陸に漂着するなどして、植物の分布が拡がることをみた。動物も同じような方法で海を渡ることは可能かもしれない。今回は海を越えた動物の移住について考えてみよう。
◎浮島に乗った漂着
前回、マダガスカルの巨鳥エピオルニスの完全な卵がオーストラリアの海岸まで流れ着いたという話をした。エピオルニスの卵の殻は厚さが3~4㎜もあるので壊れないで漂着することは可能かもしれないが、それでも生きた卵が長期間の漂流に耐えられるとは考えにくい。
動物の漂着の方法としていちばん可能性が高いのは、生きた成体が海を渡ることであるが、漂流中の食糧の問題を考えると、泳ぐことのできない動物の漂着が成功するためには、植物の果実よりも一段と高い障壁がある。
流木などに乗って漂流することも可能だが、漂流中の食糧問題がネックになるのだ。そこで海を渡るための乗り物として注目されるのが浮島である(1)。冬眠をする動物であれば、食糧なしで長期間の漂流に耐えることも可能かもしれないが、それでも流木よりは浮島のほうが乗り物としては適しているだろう。
浮島は図4-1aのように日本でも尾瀬湿原の池塘など各地で見られる。これは固定した島ではなく、枯れた草が絡まりあってできているもので、名前から分かるように水に浮いている(図4-1b)。従って、洪水などで内陸の湖でできた浮島が海に流れ出てくることもある。


たまたまそのような浮島に乗ってしまった動物が海を漂流することになるわけだが、たいていは新天地にたどり着く前に食糧が尽きるなどして死んでしまったであろう。ところが、自然はこのような試行錯誤を延々と繰り返してきたのである。
例えば10年に一度の大雨で動物を載せた大きな浮島が海に流出したとする。このようなことが100万年にわたって繰り返されたとすると、10万回の漂流があったことになる。このようなたくさんの試行の中の1回でも新天地への漂着に成功したならば、その後の進化の歴史は大きく変わることになる。
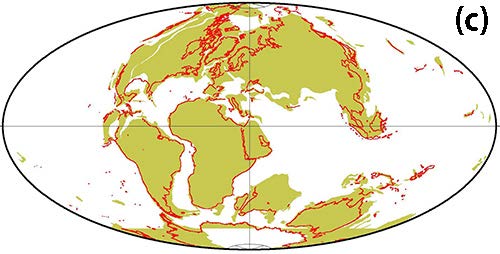
このようなことは、動物進化の歴史上何回も起ったようである。その中で最もびっくりするような移住が、およそ3500万年前に起ったと考えられるアフリカから南アメリカへの新世界ザルの祖先の移住であった。
◎新世界ザルの起源
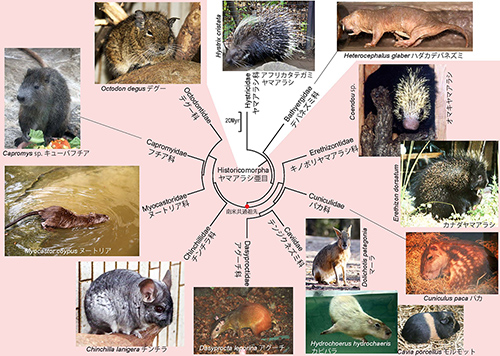
新世界の真猿類は広鼻猿類とも呼ばれている。これに対して旧世界、つまりアフリカとユーラシアの真猿類が狭鼻猿類である。ヒトもこの仲間である。図4-2は、現生の広鼻猿類の系統樹マンダラである。
南アメリカでは霊長類の古い化石は見つからないので、どこかほかの大陸からやって来たはずである。最初は広鼻猿類の祖先は北アメリカからやって来たのではないかと考えられた。北アメリカと南アメリカとはおよそ300万年前にパナマ地峡で陸続きになるまでは、離れた大陸であった。南アメリカにいちばん近い大陸は、北アメリカと南極であるが、広鼻猿類が進化した頃には南極はすでに氷の大陸になっており、南極大陸経由は考えられない。
一方、北アメリカは離れているとはいっても、そこから漂着などでたどり着いた可能性はあるだろう。ダーウィンがビーグル号で南アメリカのパタゴニアを調査した際に、その化石を発見したマクラオケニアという絶滅哺乳類がいた。ダーウィンはこの動物がラクダのような長い首をもった巨獣であると表現しており、同じく南アメリカに生息するグアナコやラマに近縁ではないかと考えたが(5)、この動物が系統的にどのようなところに位置するかは不明であった。
この動物は滑距(かっきょ)目(Litopterna)に分類されているが、およそ700万年前~2万年前に南アメリカに生息していた。最近、絶滅動物のDNAを調べる古代DNA解析により、この動物がウマ、サイ、バクなど奇蹄目に近縁であることが示された(6)。
メスが胎盤をもった哺乳類である真獣類の中で、食肉目(イヌ、クマ、ネコ)、鱗甲目(センザンコウ)、奇蹄目(ウマ、サイ、バク)は系統的にまとまったグループを作る。この中には、イヌ、ネコ、ウマなど、ヒトが昔から家畜化して友達として接してきた動物が多く含まれることから、友獣類(Zooamata)という(8)。Zooamataはギリシャ語の動物(zoo)とラテン語の友達(amata)からきている。図4-3がマクラオケニアを含めた友獣類の系統樹マンダラであるが、マクラオケニアは奇蹄目に属することが分かる。友獣類や鯨偶蹄目はもともと北半球の大陸で進化したものであり、南アメリカには生息していなかった。
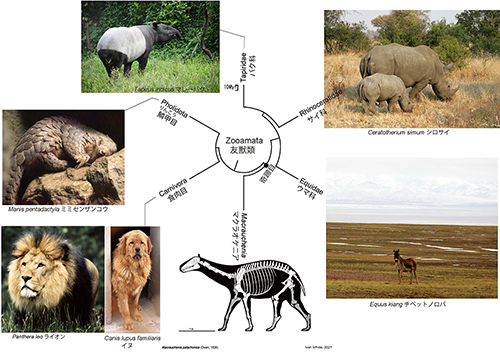
ところが同じように広鼻猿類の祖先が北アメリカからやって来たと考えることはできないのだ。なぜならば、北アメリカからは広鼻猿類の祖先になりそうな霊長類の化石がまったく見つからないのである。
一方、南アメリカからはるかに離れたアフリカのエジプトで、広鼻猿類の祖先になり得ると思われる霊長類の化石が見つかったのだ。そのようなことから、広鼻猿類の祖先はアフリカから大西洋を渡って南アメリカにやって来たものに違いないと考えられるようになったのである。これはとんでもなくありそうもないことに思われるかもしれないが、そのようなことが起こる蓋然性を高める要素が2つある。
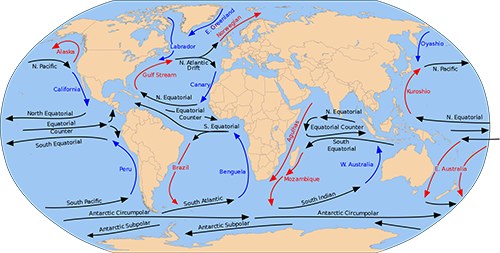
一つはアフリカと南アメリカを隔てる距離が、およそ3500万年前は現在よりも短かったということである。ゴンドワナ超大陸分裂の一環として、およそ1億500万年前にアフリカと南アメリカが分かれて、大西洋が生まれた。その後大西洋がだんだんと広がって、現在のようになるわけだが、広鼻猿類の祖先がおよそ3500万年前にアフリカから南アメリカに渡って来たのだとすると、その頃の2つの大陸の距離は現在の半分程度だったと考えられる(図4-4)。
次に重要なのは海流の方向である。前回の図3-3に現在の世界の主要な海流を示した。
ところで、マクラオケニアなど南アメリカの多くの絶滅哺乳類の化石を発見したダーウィンは、『ビーグル号航海記』の中で次のように述べている:
近年、ルントとクラウゼン両氏がブラジルの洞窟からヨーロッパに運びこんだ大コレクションである。…この絶滅種数は、現生の種数よりもずっと多い。化石種のアリクイ、アルマジロ、バク、ペッカリー、グアナコ、オポッサム、数多くの南アメリカ産齧歯類とサル類、そのほかである。同一の大陸で見られる絶滅種と現生種にかかわるこの驚くべきつながりは、将来、地球の生物の出現と消滅の問題に対し、ほかのどのような事実資料よりも多くの光を投げかけてくれるものと、わたくしは確信している。
ダーウィンは1859年に『種の起源』を出版して彼の進化理論を公表するよりもはるか以前から種の間の進化的なつながりをはっきりと意識していたのである。
新世界ザルの祖先がアフリカからやってきたことを直接証明する証拠はないが、現在のところそれ以外に考えようがないのだ。あり得ないことを取り除いていって残った可能性は、たとえそれが信じ難いことに見えても受け入れざるを得ない。マクラオケニアの祖先が北アメリカから海を渡ってやってきたように、新世界ザルの祖先はもっと遠いアフリカから同じく海を渡ってやって来たのである。
【引用文献】
1. Van Duzer, C. (2004) “Floating Islands”. Cantor Press.
2.長谷川政美(2011)『新図説・動物の起源と進化』八坂書房.
3.Kiesling, N.M.J., Yi, S.V., Xu, K., et al. (2015) The tempo and mode of New World monkey evolution and biogeography in the context of phylogenomic analysis. Mol. Phylogenet. Evol. 82B, 386-399.
4.Kierulff, M.C.M., Ruiz-Miranda, C.R., de Oliveira, P.P., et al. (2012) The Golden lion tamarin Leontopithecus rosalia: a conservation success story. Int. Zoo Yb. 46, 36-45.
5.チャールズ・R・ダーウィン(1845)『新訳・ビーグル号航海記』(荒俣宏訳、訳本は2013年、平凡社).
6.Westbury, M., Baleka, S., Barlow, A., et al. (2017) A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nature Comm. 8, 15951.
7.dos Reis, M., Inoue, J., Hasegawa, M., et al. (2012) Phylogenomic datasets provide both precision and accuracy in estimating the timescale of placental mammal phylogeny. Proc. Roy. Soc. London B. 279, 3491–3500.
8.Waddell, P.J., Okada, N., Hasegawa, M. (1999) Towards resolving the interordinal relationships of placental mammals. Syst. Biol. 48, 1-5.
9.Fabre, P.-H., Hautier, L., Dimitrov, D., et al. (2012) A glimpse on the pattern of rodent diversification: a phylogenetic approach. BMC Evol. Biol. 12, 88.
*もっと「進化」について知りたい人の入門書:
☆本連載が本になりました!
長谷川政美著『進化生物学者、身近な生きものの起源をたどる
』 (ベレ出版)。 イヌやネコやクマなど身近な生き物はすべて進化していまここにいる。もちろんヒトも。生き物の進化が一目でわかる「系統樹マンダラ」を多数掲載(系統樹の「見方」付き)。ささやかな「現代版 種の起原」ができました。

☆はじめの一冊にオススメ!
長谷川政美監修・畠山泰英構成『世界でいちばん素敵な進化の教室
』 (三才ブックス)。 本書は美しい写真とQ&A形式の簡潔な文章で、38億年におよぶヒトを含む生き物の進化を解説した超入門ビュアルブックです。子供から大人まで気軽に楽しんでいただけます。
4刷(2022年10月)。





{kind=link}